Embed Size (px)
Citation preview

ORIGINAL PAPER
Adaptations to captive breeding of the longhorn beetle Morimusfunereus (Coleoptera: Cerambycidae); application on amylasestudy
Biljana Dojnov • Zoran Vujcic • Natasa Bozic •
Aleksandra Margetic • Miroslava Vujcic •
Vera Nenadovic • Jelisaveta Ivanovic
Received: 18 June 2010 / Accepted: 2 June 2011 / Published online: 23 June 2011
� Springer Science+Business Media B.V. 2011
Abstract Captive breeding has been suggested as a
method of conservation for many vertebrates, and is
increasingly being proposed as a strategy for invertebrates.
In this study, the growth, development and fertility of
adults of the vulnerable cerambycid Morimus funereus
reared in captivity are examined. Two oviposition cycles;
from May to September and from January to March were
studied and larvae from wild adults and from the progeny
of captive adults (second generation larvae) were exam-
ined. Five to 12 instars were observed during larval
development. Larval development was completed in
218 days (average) for the progeny of wild adults with an
average mortality rate of 10.3% and in 226 days (average)
for larvae from captive adults with mortality rate of 34.9%.
First generation larval body weights were disparate during
development, while second generation larvae had similar
weights with no significant differences. In this study we
have tested the potential of captive breaded M. funereus
larvae as a model for investigation of digestive enzymes.
Amylase from the midgut of larvae reared under laboratory
conditions showed twofold higher specific activities with a
decreased number of isoforms expressed, as compared to
the enzyme from field-collected larvae. Captive breeding
of M. funereus can be used in the future as a part of
an effective conservation strategy for this rare insect
species.
Keywords Cerambycidae � Morimus funereus �Larval development � Midgut a-amylase � Isoform
Introduction
Morimus funereus (Coleoptera: Cerambycidae) inhabits a
relatively narrow geographical zone (forests of south-
eastern Europe), and infests deciduous and coniferous trees
(IUCN 2009). This cerambycid beetle develops on host
plants of the families Tiliaceae, Fagaceae, Corylaceae,
Salicaceae, Fabaceae and Pinaceae, using either physio-
logically debilitated trees, tree stumps or recently cut logs.
The larvae are inner bark (phloem) feeders, found in the
first stage of decomposition of trees (Ivanovic 1968). There
are only a few records of development and longevity in
wildlife. Larval development of M. funereus lasts
3–4 years (Council of Europe 1996). Adults have a rela-
tively long duration of about 2 years (Council of Europe
1996; Vrezec et al. 2010), and are able to slowly disperse
over this period, despite being flightless.
The taxonomy and nomenclature of species of the genus
Morimus has alternative interpretations. Morimus funereus
and Morimus ganglbaueri are often considered as syn-
onyms of Morimus asper. According to Simoneta (1989)
all are good species but Sama (1988) describes M. funereus
and M. ganglbaueri as subspecies of M. asper. Preliminary
results using molecular data suggest that the European
Morimus actually represents a single, genetically and
morphologically variable species (Antonini et al. 2010).
B. Dojnov � N. Bozic � A. Margetic � M. Vujcic
Center of Chemistry, Institute of Chemistry, Technology and
Metallurgy, University of Belgrade, Studentski trg 12-16,
Belgrade, Serbia
Z. Vujcic (&)
Department of Biochemistry, Faculty of Chemistry, University
of Belgrade, Studentski trg 12-16, 11000 Belgrade, Serbia
e-mail: [email protected]
V. Nenadovic � J. Ivanovic
Department of Insect Physiology and Biochemistry, Institute for
Biological Research Sinisa Stankovic, Despota Stefana 142,
Belgrade, Serbia
123
J Insect Conserv (2012) 16:239–247
DOI 10.1007/s10841-011-9411-x

M. funereus is listed as vulnerable by the IUCN Red list
of threatened species (IUCN 2009) and is an Annex II
Species of the EU Habitats and Species Directive (2009)
and also by the Decree on designate and protect the strictly
protected and endangered species of wild plants, animals
and fungi (2010). The number of individuals within pop-
ulation of M. funereus and the distribution of populations
are not well understood. Romero-Samper and Bahillo
(1993) describe the distribution of M. funereus and
M. asper in the Mediterranean and Adriatic region
including Italy but detailed assessments for the Pannonian
and Mediterranean regions remain to be performed. Data
are available for the Italian region and they are included in
Annex II Species of the EU Habitats and Species Directive.
Projects have been started in Italy and Slovenia in 2009 to
provide guidelines for monitoring and conservation of
saproxylic beetles (including M. funereus) including the
use of standard methodologies for measuring population
stabilities and trends, as well as habitat ranges (Campanaro
et al. 2010; Ambrozic et al. 2010). It is expected that more
information will become available on the distribution and
abundance of M. funereus in the future.
M. funereus had been found at 40 locations in Serbia in
1965 (Adamovic 1965) but are now decreasing, recently
being recorded from only 17 locations (Ilic 2005). In one
nature reserve in Serbia only five M. funereus adults were
found during the period between 2000 and 2004 (Pil and
Stankovic 2006).
The decline of M. funereus results from a combination
of factors. Larger dead trees, on which it dependent, have
been removed from numerous forest ecosystems in the
past. M. funereus adults are flightless with peak activity
between 8 p.m. and 3 a.m. in the summer time—in the first
half of May, and in the second half of June. (Vrezec et al.
2010). Low mobility of these insects in combination with
continued loss of resources has brought about a reduction
of M. funereus and the numbers can be expected to
decrease in the future.
Captive breeding is increasingly being suggested as a
valuable and cost-effective conservation tool for certain
invertebrates (Pearce-Kelly et al. 1998; Lewis and Thomas
2001). Conservation management of M. funereus is cur-
rently focused on monitoring in the field and there are no
existing programs for active conservation in Serbia. The
lack of opportunities for conservation management of
M. funereus is a strong justification for investment in
captive rearing. In many ways captive breeding and
resource restoration have to go hand in hand.
M. funereus is a also a good model for the study of the
impacts of various types of stress on insect metabolism
(Ivanovic et al. 1975; Stanic et al. 1985; Nenadovic et al.
1986; Ivanovic 1991; Jankovic-Hladni et al. 1992; Ðordevic
et al. 1999; Lekovic et al. 2001) and for purification and
characterization of digestive enzymes (Bozic et al. 2008;
Dojnov et al. 2008; Loncar et al. 2010). Recent papers sug-
gest great plasticity of M. funereus larvae, therefore, it is
anticipated that M. funereus can be captively bred.
The amount and isoform production of digestive
enzymes in captive reared larvae can be a useful indicator
of how captive populations respond to captivity. Since it
has been purified and characterized from the midgut of
larvae developed in the wild (Dojnov et al. 2008),
a-amylase can be chosen as a model enzyme for how
M. funereus succeed in captivity. The possibility of using
captive larvae for purification of digestive enzymes could
enables retrieval of sufficient quantities of source material
without disturbance of the wildlife population.
The aims of this work are to examine the feasibility of
captively breeding M. funereus and to assess whether the
administered diet results in reproductive M. funereus adults
and to describe larval development in captivity.
Materials and methods
All reagents and solvents used were purchased from Merck
(Darmstadt, Germany) and Sigma–Aldrich (St. Louis, MO,
USA), unless otherwise stated. ‘‘Palenta’’ (‘‘Mitrosrem’’,
Sremska Mitrovica, Serbia) was produced from corn grits.
Two groups of M. funereus adults were used in this
study: adults, collected from oak stumps on Fruska Gora
Mountain during spring and then reared under laboratory
conditions (wild adults); and captive adult progeny
obtained by breeding from these wild adults for a period of
3 years.
Three groups of M. funereus larvae were examined in
this study: larvae collected on Fruska Gora Mountain
during spring and dissected upon collection, and first and
second generation reared larvae. A detailed description of
each adult and larval group is presented in Table 1.
For each year of this study, the number of adults and
larvae taken from the field was confined to the number
proposed in permits to work with protected species,
obtained from the relevant institutions; Institute for Nature
Conservation of Serbia and Ministry of Environment and
Spatial Planning, Serbia.
One female and one or two males from the same adult
group were put in a transparent plastic container
(18 9 12 9 12 cm) with holes in the lid for ventilation.
Oak leaves and bark placed in the containers covered with
filter paper, served as food, hiding places and oviposition
site. Seasonal fruits, lucerne, and crumbled sweet crackers
were added as additional food. Water was supplied by use
of wet cotton wool. The relative humidity (RH) in the
laboratory was about 50%. The average annual tempera-
tures varied between 22�C in the cold and 26�C in the hot
240 J Insect Conserv (2012) 16:239–247
123

season. Leaves and other food were replaced with fresh
material two times a week. Deposited eggs were collected
daily, placed on dietary media in Petri dishes and trans-
ferred to 23�C and RH of about 50%. Eggs were examined
on a daily basis for hatching.
M. funereus larvae were reared under laboratory condi-
tions from eggs to adults. The artificial diet used was com-
posed of (g/L): ‘‘Palenta’’, 100; agar–agar, 10; sugar, 25; dry
Brewer’s yeast, 25. Diet (medium) was prepared by baking.
Nipagin (methyl ester-p-hidroxybenzoic acid) (0.2 g/mL
ethanol) was added to the medium after cooling to below
70�C. Warm media was plated out on round–bottom plastic
boxes. The chemical composition of commercial polenta
‘‘Palenta’’ is: water 13–15%, fat 0.5–1.5%, cellulose
0.35–0.70%, proteins 7–9%, soluble starch 70–80% and ash
0.4–0.8%. Raw starch in corn was converted to digestible
dextrin chains by swelling, heating and mechanical grinding.
Due to the cannibalistic behaviour of M. funereus larvae,
individual larvae were each placed separately in a round-
bottom plastic box (diameter 6 cm) with ventilation holes on
top. During development, larvae were kept in the dark at
23�C and RH 50%. Fresh dietary medium was given and
replaced with fresh one and weight of larvae weighed each
week. Larvae were examined daily for molting.
Three different crude midgut extracts were prepared
from wild larvae and first and second generation captive
larvae. Wild larvae had body weights ranging from 1.0 to
1.5 g, and mesenteron weights ranging from 0.11 to 0.33 g.
Captive larvae had body weights ranging from 2.3 to 3.2 g,
and mesenteron weights ranging from 0.22 to 0.50 g. Lar-
vae were dissected during the sixth instar, when they were
feeding actively. After decapitation, the midguts were
isolated on ice, weighed and homogenized, using a pre-
chilled mortar and pestle, in 2 vol. (g/mL) of ice-cold 0.9%
NaCl in 0.1 M pH 5 acetate buffers with addition of quartz
powder. The crude extracts were prepared as described by
Bozic et al. (2003).
a-Amylase activity was assayed using the dinitrosali-
cylic acid (DNS) procedure of Bernfeld (1955) with use of
soluble starch as substrate. Samples (50 lL) were incu-
bated in 450 lL 50 mM acetate buffer pH 5.0 containing
1.0% (w/v) starch, 2.0 mM NaCl and 0.1 mM CaCl2 at
35�C for 30 min. Maltose was used as standard. Each data
point represents the mean of three independent assays
(standard errors were less than 5% of the means). One unit
of a-amylase activity was defined as the amount of enzyme
required to produce 1 lmol maltose in 1 min at 35�C.
Midgut a-amylases were detected using in-gel activity
staining following native PAGE and isoelectric focusing
(IEF) following Dojnov et al. (2008). IEF was performed
using the Multiphor II electrophoresis system (Pharmacia-
LKB Biotechnology), on 7.5% polyacrylamide gel with
ampholytes in the 2.5–4.5 pH range. Low pI kit (GE
Healthcare) was used to provide pI markers.
Protein concentrations were determined by the Bradford
method (Bradford 1976) using bovine serum albumin as the
protein standard. Each data point represents the mean of
three independent assays ± SEM (standard errors were
less than 5% of the means). Proteins extracted from mid-
guts were examined by SDS PAGE according to Laemmli
(1970). Low molecular weights—SDS marker kit (GE
Healthcare) was used as molecular mass standards. After
electrophoresis the gel was stained with Coomassie bril-
liant blue (CBB).
For statistical analysis GraphPad Prism 5 program was
used. One–way analysis of variance (ANOVA) with Tukey
post-hoc tests was used to test for differences in egg laying
and larval weights between groups. The number of days for
each instar during larval development was analyzed using
the D’Agostino and Pearson omnibus normality test. The
durations of instars for the two captive bred larval groups
were compared using t tests with 95% confidence intervals.
Results
Behavior and feeding of M. funereus adults reared
in captivity
The activity of adults reared in the laboratory depended on
the season and environmental factors. During spring and
summer adults fed actively on seasonal fruit, lucerne stems,
Table 1 Description of terms used in the paper for the examined groups of larvae and adults
Denotation Description Number
Wild adults Adults, collected from the natural habitat 30
Captive
adults
Adults obtained in captivity from first generation of captive larvae 31
Wild larvae Larvae collected in nature from oak stumps 44
Captive
larvae I
First generation of captive larvae—larvae obtained from eggs deposited by wild adults and
reared in the laboratory
507, of which 31 were reared
to adults
Captive
larvae II
Second generation of captive larvae—larvae obtained from eggs deposited by captive adults 73, of which 30 were reared
to adults
J Insect Conserv (2012) 16:239–247 241
123

crackers and fresh oak bark and leave and mated repeat-
edly. But during autumn and winter, adults were clustered
on the bark or hidden beneath leaves with minimal activity.
During that time adults consumed only seasonal oak bark
and leaves collected in the field. Nevertheless, some adults
of both groups engaged in active feeding and in January.
Oviposition
Oval, elongated 3–4 mm long eggs, protected by rigid
chorine, were found on bark, leaves, filter paper, on the
bottom of the container or inserted into bark crevices. Two
oviposition cycles were detected under our laboratory
conditions, from May to September and from January to
March. Females usually deposited 1 or 2 eggs per day and
the number of eggs deposited by each female differed
(Table 2). The number of males presented to the females in
a box (1 or 2 males), did not affect the number of eggs
deposited. Wild females from the field deposited more eggs
than captive females. More eggs were found during the first
oviposition cycle in both groups of females. Difference
between oviposition cycles in two examined groups were
analyzed by ANOVA test (F3,45 = 5.525, P = 0.0026).
Tukey’s Multiple Comparison Test show a significant
statistical difference between the first and second oviposi-
tion cycles in the group of females from the nature
(**P \ 0.05) and also show statistical difference between
first cycles of two groups (*P \ 0.05) and between first
cycle in females from nature and second cycle in captive
females (*P \ 0.05). 17.65% of females from the wild and
22.23% of females in captivity were not carrying eggs.
80.98% of eggs from wild adults were viable and
72.67% produced by captive adults were viable, but there
were no significant differences in viability between groups
and within groups (P [ 0.10 in both cases).
Larval development
Larval development durations and number of instars were
variable. Larval development varied between five and
twelve with most larvae having five or six instars prior to
pupation. In general, males developed with fewer (five)
instars prior to pupation than females (six or more instars).
Average larval development of faster developing individ-
uals (subgroup I) was completed after 218 days in the first
generation of larvae, and after 226 days in the second
generation. 52% of larvae from the first generation and
21% of larvae from the second generation took to develop
Table 2 Number of deposited eggs per M. funereus female during two oviposition cycles for wild and captive adults
No of female Wild adults Captive adults
First oviposition cycle Second oviposition cycle First oviposition cycle Second oviposition cycle
Total of eggs/female Total of eggs/female Total of eggs/female Total of eggs/female
1 12 0 0 13
2 14 7 17 24
3 4 0 20 7
4 22 10 4 0
5 38 6 4 0
6 53 0 0 0
7 3 0 4 8
8 9 / 31 4
9 0 / 0
10 54 0
11 51 15
12 59 10
13 47 38
14 29 2
15 82 /
16 0 /
17 0 0 /
Total 477aaa 88baa 80caa 56daa
Results were statistically analyzed with one-way ANOVA using Tukey test (a = 0.05), P = 0.0026, F = 5.525, r2 = 0.2692. Oviposition cycles
I and II, of both wild adults and captive adults, were compared within one and between both groups of larvae. Values followed by the same letter
in columns are not significantly different
242 J Insect Conserv (2012) 16:239–247
123

from the last molting to pupation (subgroup II), and
development to adults lasted on average 278 days and
313 days, respectively (Table 3).
There were significant differences between first and
second generation captive larvae in terms of time needed
for egg hatch, sixth instar duration and period from last
molting to pupation (Table 3). Other instars were similar
for first generation larvae and second generation larvae.
Significant differences were also obtained within both
groups for development during the third instar, according
to the D’Agostino and Pearson omnibus normality test
(P \ 0.0001). The duration of the pupal stage period was
similar in both groups.
On adult emergence a sex ratio of 1:1 (F:M) was
obtained from first generation larvae, and 1:1.2 (F:M) from
second generation individuals. These were not significantly
different (Fisher’s exact test P & 1.0).
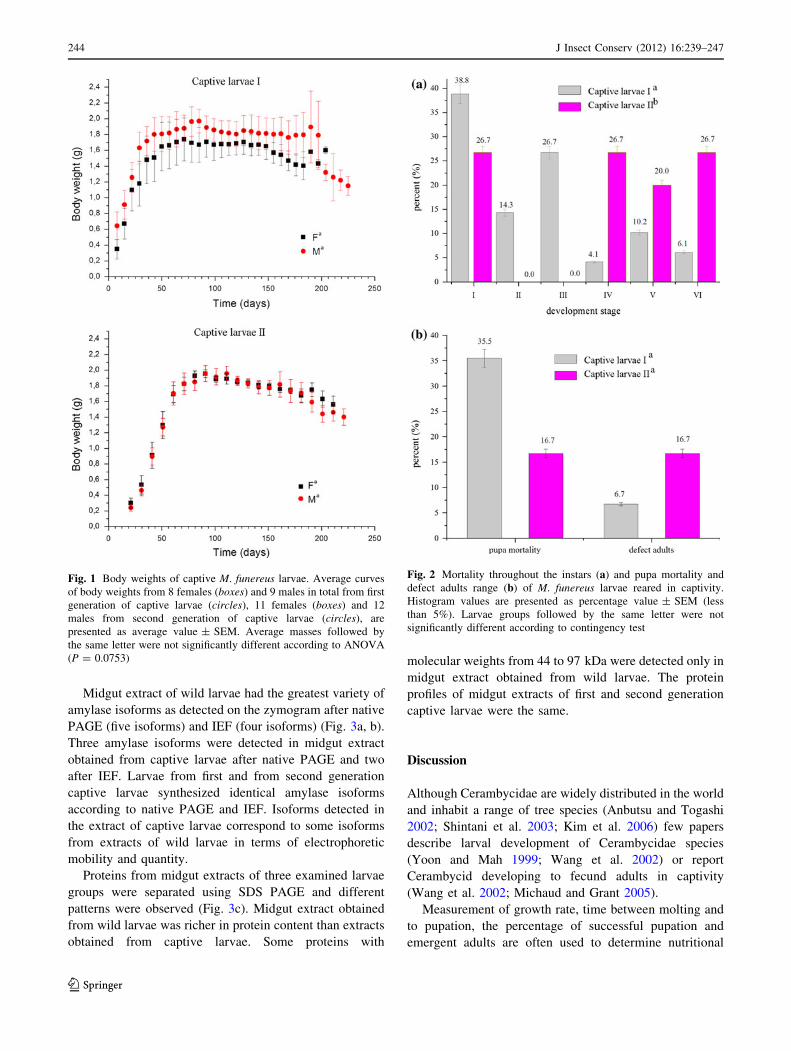
Average larval weight curves are presented in Fig. 1.
Females and males from second generation larvae had sim-
ilar weights during development and there were no signifi-
cant differences between or within the groups (males
ANOVA results: F11,155 = 1.574, P = 0.1115; females
ANOVA results F10,151 = 0.6262, P = 0.7897). Females
attained a maximum weight of 1.94 g and males 1.96 g. Loss
of body mass was observed in the days before and during
molting, and in days prior to pupation in the second gener-
ation. By contrast, body weights of females and males from
the first generation larvae were disparate. With significant
differences being observed within female and male larval
groups (males ANOVA results: F8,201 = 34.61,
P \ 0.0001***; females ANOVA results F8,58 = 13.58,
P \ 0.0001***). Females of the first generation needed less
time to achieve maximum body weight (1.75 g) than males
(1.96 g). Prior to entering the pupal stage females lost more
weight than males. There were no significant statistical
differences between the weights of first generation and sec-
ond generation during develop in captivity, neither for males
or for females (F3,98 = 1.010, P = 0.3919).
Mortality rates in both examined groups of captive lar-
vae, intended for dissection in the sixth developmental
stage, are shown in Fig. 2a. Most first generation mortality
was during the first and third instars, while most of second
generation larvae died during the first, fourth and sixth
instars. There were significant statistical differences
(P \ 0.05) in the mortality of these two larval groups
through all larval instars (v2 = 18.15, df = 5, P = 0.0028).
Overall mortality in the first generation larvae was 9.7 and
20.6%) in the second generation.
In the first generation, 16.7% of larvae developed into
defective adults, while this percentage was 6.7% in the
second generation (Fig. 2b). Defective adults were not able
to eat or move and they died soon after metamorphosis. It
was noted that all defective adults were larvae with
extended period of the last larval instar (subgroup II,
Table 2). There were no significant statistical differences
(P \ 0.05) in the combined number of defective adults and
pupal mortality of the two groups (Fisher’s exact test,
P = 0.169).
Amylase activity and protein concentration
in midgut extracts
The activity of amylase (62 U/mL) and concentration of
proteins (11 mg/mL) were higher in the crude midgut
extract of M. funereus larvae collected from the field as
compared to extracts obtained from larvae developed in the
laboratory (50 U/mL; 4–5 mg of protein/mL), (Table 4).
However, specific amylase activities (U/mg) were twofold
higher in midgut extracts of captive larvae than in midguts
obtained from field larvae.
Table 3 Duration of developmental stages of M. funereus larvae reared in the laboratory, expressed in number of days
Developmental stage Captive larvae I Captive larvae II
Mean ± SEM N Mean ± SEM N P
Egg to larva 11.00 ± 0.49 31 12.73 ± 0.58 30 0.0271
First instar 8.45 ± 0.53 31 10.47 ± 1.04 30 N.S.
Second instar 11.31 ± 1.05 29 13.73 ± 1.58 30 N.S.
Third instar 15.52 ± 2.27 27 14.86 ± 1.63 28 N.S.
Fourth instar 23.26 ± 4.09 27 20.96 ± 1.43 25 N.S.
Fifth instar 31.00 ± 7.24 22 25.73 ± 1.68 15 N.S.
Sixth instar 22.55 ± 3.36 11 50.33 ± 6.44 3 0.0024
Last molting to pupation (subgroup I) 72.89 ± 4.27 9 54.77 ± 2.57 22 0.0008
Last molting to pupation (subgroup II) 132.30 ± 6.30 12 141.80 ± 6.18 6 N.S.
Pupa to adult 21.88 ± 0.84 17 22.14 ± 1.38 22 N.S
P = Significance of t test between cative larve groups, N.S. = P [ 0.05
J Insect Conserv (2012) 16:239–247 243
123
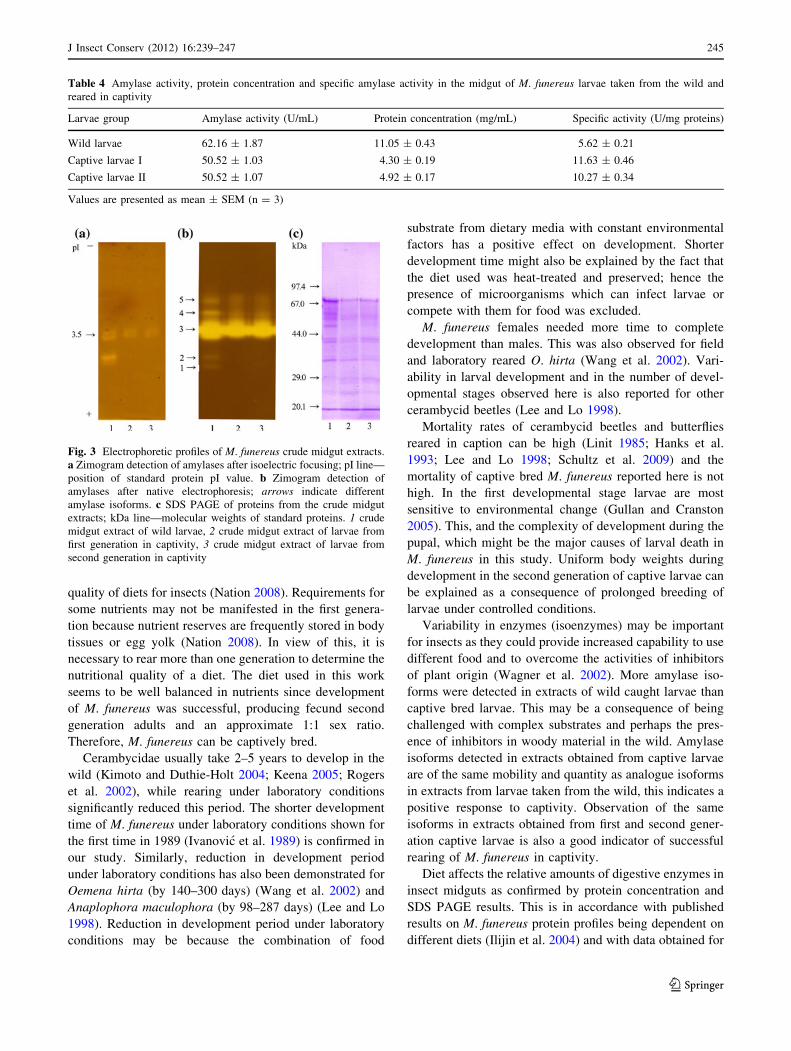
Midgut extract of wild larvae had the greatest variety of
amylase isoforms as detected on the zymogram after native
PAGE (five isoforms) and IEF (four isoforms) (Fig. 3a, b).
Three amylase isoforms were detected in midgut extract
obtained from captive larvae after native PAGE and two
after IEF. Larvae from first and from second generation
captive larvae synthesized identical amylase isoforms
according to native PAGE and IEF. Isoforms detected in
the extract of captive larvae correspond to some isoforms
from extracts of wild larvae in terms of electrophoretic
mobility and quantity.
Proteins from midgut extracts of three examined larvae
groups were separated using SDS PAGE and different
patterns were observed (Fig. 3c). Midgut extract obtained
from wild larvae was richer in protein content than extracts
obtained from captive larvae. Some proteins with
molecular weights from 44 to 97 kDa were detected only in
midgut extract obtained from wild larvae. The protein
profiles of midgut extracts of first and second generation
captive larvae were the same.
Discussion
Although Cerambycidae are widely distributed in the world
and inhabit a range of tree species (Anbutsu and Togashi
2002; Shintani et al. 2003; Kim et al. 2006) few papers
describe larval development of Cerambycidae species
(Yoon and Mah 1999; Wang et al. 2002) or report
Cerambycid developing to fecund adults in captivity
(Wang et al. 2002; Michaud and Grant 2005).
Measurement of growth rate, time between molting and
to pupation, the percentage of successful pupation and
emergent adults are often used to determine nutritional
Fig. 1 Body weights of captive M. funereus larvae. Average curves
of body weights from 8 females (boxes) and 9 males in total from first
generation of captive larvae (circles), 11 females (boxes) and 12
males from second generation of captive larvae (circles), are
presented as average value ± SEM. Average masses followed by
the same letter were not significantly different according to ANOVA
(P = 0.0753)
Fig. 2 Mortality throughout the instars (a) and pupa mortality and
defect adults range (b) of M. funereus larvae reared in captivity.
Histogram values are presented as percentage value ± SEM (less
than 5%). Larvae groups followed by the same letter were not
significantly different according to contingency test
244 J Insect Conserv (2012) 16:239–247
123
quality of diets for insects (Nation 2008). Requirements for
some nutrients may not be manifested in the first genera-
tion because nutrient reserves are frequently stored in body
tissues or egg yolk (Nation 2008). In view of this, it is
necessary to rear more than one generation to determine the
nutritional quality of a diet. The diet used in this work
seems to be well balanced in nutrients since development
of M. funereus was successful, producing fecund second
generation adults and an approximate 1:1 sex ratio.
Therefore, M. funereus can be captively bred.
Cerambycidae usually take 2–5 years to develop in the
wild (Kimoto and Duthie-Holt 2004; Keena 2005; Rogers
et al. 2002), while rearing under laboratory conditions
significantly reduced this period. The shorter development
time of M. funereus under laboratory conditions shown for
the first time in 1989 (Ivanovic et al. 1989) is confirmed in
our study. Similarly, reduction in development period
under laboratory conditions has also been demonstrated for
Oemena hirta (by 140–300 days) (Wang et al. 2002) and
Anaplophora maculophora (by 98–287 days) (Lee and Lo
1998). Reduction in development period under laboratory
conditions may be because the combination of food
substrate from dietary media with constant environmental
factors has a positive effect on development. Shorter
development time might also be explained by the fact that
the diet used was heat-treated and preserved; hence the
presence of microorganisms which can infect larvae or
compete with them for food was excluded.
M. funereus females needed more time to complete
development than males. This was also observed for field
and laboratory reared O. hirta (Wang et al. 2002). Vari-
ability in larval development and in the number of devel-
opmental stages observed here is also reported for other
cerambycid beetles (Lee and Lo 1998).
Mortality rates of cerambycid beetles and butterflies
reared in caption can be high (Linit 1985; Hanks et al.
1993; Lee and Lo 1998; Schultz et al. 2009) and the
mortality of captive bred M. funereus reported here is not
high. In the first developmental stage larvae are most
sensitive to environmental change (Gullan and Cranston
2005). This, and the complexity of development during the
pupal, which might be the major causes of larval death in
M. funereus in this study. Uniform body weights during
development in the second generation of captive larvae can
be explained as a consequence of prolonged breeding of
larvae under controlled conditions.
Variability in enzymes (isoenzymes) may be important
for insects as they could provide increased capability to use
different food and to overcome the activities of inhibitors
of plant origin (Wagner et al. 2002). More amylase iso-
forms were detected in extracts of wild caught larvae than
captive bred larvae. This may be a consequence of being
challenged with complex substrates and perhaps the pres-
ence of inhibitors in woody material in the wild. Amylase
isoforms detected in extracts obtained from captive larvae
are of the same mobility and quantity as analogue isoforms
in extracts from larvae taken from the wild, this indicates a
positive response to captivity. Observation of the same
isoforms in extracts obtained from first and second gener-
ation captive larvae is also a good indicator of successful
rearing of M. funereus in captivity.
Diet affects the relative amounts of digestive enzymes in
insect midguts as confirmed by protein concentration and
SDS PAGE results. This is in accordance with published
results on M. funereus protein profiles being dependent on
different diets (Ilijin et al. 2004) and with data obtained for
Table 4 Amylase activity, protein concentration and specific amylase activity in the midgut of M. funereus larvae taken from the wild and
reared in captivity
Larvae group Amylase activity (U/mL) Protein concentration (mg/mL) Specific activity (U/mg proteins)
Wild larvae 62.16 ± 1.87 11.05 ± 0.43 5.62 ± 0.21
Captive larvae I 50.52 ± 1.03 4.30 ± 0.19 11.63 ± 0.46
Captive larvae II 50.52 ± 1.07 4.92 ± 0.17 10.27 ± 0.34
Values are presented as mean ± SEM (n = 3)
Fig. 3 Electrophoretic profiles of M. funereus crude midgut extracts.
a Zimogram detection of amylases after isoelectric focusing; pI line—
position of standard protein pI value. b Zimogram detection of
amylases after native electrophoresis; arrows indicate different
amylase isoforms. c SDS PAGE of proteins from the crude midgut
extracts; kDa line—molecular weights of standard proteins. 1 crude
midgut extract of wild larvae, 2 crude midgut extract of larvae from
first generation in captivity, 3 crude midgut extract of larvae from
second generation in captivity
J Insect Conserv (2012) 16:239–247 245
123

other insects (Terra and Ferreira 1994). Trypsin-like
enzymes were present only in the midgut of M. funereus
larvae reared under laboratory conditions using this artifi-
cial diet (Loncar et al. 2009), while they were not present
in field larvae (Bozic et al. 2003).
Plasticity of M. funereus has been well studied (Ivanovic
1970) and is confirmed in this study by modification of
developmental period, number of larval instars, and vari-
ability in enzymes (isoforms). Overall we have shown that
M. funereus is suitable for captive breeding. However, the
range of amylase isoforms expressed in captive bred indi-
viduals is less than in wild caught individuals.
A breeding program of M. funereus may help in the
conservation of this rare beetle if combined with landscape
and forest management. Conservation management should
consist of preservation of resources for M. funereus, estab-
lishment of standard methodologies for measurement of
population sizes and trends, field investigation on phenology
and behaviour, captive breeding and, finally, reintroduction
of adults into the wild. Captive bred individuals can be used
to supplement existing populations rather than for estab-
lishing new populations, due to the fact that local environ-
mental factors, that can pose a threat to these insects, are still
to be determined. If relevant data on the behaviour and
location of adults in the wild is obtained at the time of capture
the choice of locations for introduction will be easier.
Progeny of the field population (first generation of captive
insects) can be introduced into the wild, because they are
fecund and provide for a satisfactory amount of viable eggs,
similarly to adults collected in field. The ability to rear
fecund second generation individuals also indicates it may be
possible to establish captive stocks for a more substantial
captive breeding program. In addition our methodology
provides an approach that may be of value in conservation
efforts focused on other Cerambycidae.
Acknowledgments This study was supported by a grant from the
Serbian Ministry of Science and Technological Development (project
grant number 172048). We are very grateful to Dejan Stojanovic and
National Park ‘‘Fruska Gora’’ for their kind assistance by collection of
M. funereus field adults used in these experiments. We are grateful to
Jasna Bojcevski and Tim Shreeve for help in translating our English.
References
Adamovic Z (1965) Cerambycidae (Coleoptera) collected in Serbia.
Glasnik Prirodnjackog muzeja srpske zemlje 20:147–183 [in
Serbian]
Ambrozic S, Kapla A, Vrezec A (2010) Monitoring scheme of
saproxylic beetles in the scope of Natura 2000 in Slovenia. Book
of abstracts, 6th European symposium on the conservation of
saproxylic beetles, Ljubljana, p 16
Anbutsu H, Togashi K (2002) Oviposition deterrence associated with
larval frass of the Japanese pine sawyer, Monochamus alternatus(Coleoptera: Cerambycidae). J Insect Physiol 48:459–465
Antonini G, Cerretti P, Trizzino M, Campanaro A, Mason F,
Biscaccianti A, Audisio P (2010) Morimus asper (Sulzer,
1776) and its Western Palaearctic allies: genetically supported
natural entities or taxonomic artefacts? Book of abstracts, 6th
European symposium on the conservation of saproxylic beetles,
Ljubljana, pp 39–40
Bernfeld P (1955) Amylases, a and b. In: De Murray P (ed) Methods
in enzymology, vol I. Deutcher Acad. Press INC, San Diego,
pp 149–158
Bozic N, Vujcic Z, Nenadovic V, Ivanovic J (2003) Partial
purification and characterization of midgut leucyl aminopepti-
dase of Morimus funereus (Coleoptera: Cerambycidae) larvae.
Comp Biochem Physiol 134B:231–234
Bozic N, Ivanovic J, Nenadovic V, Bergstroem J, Larsson T, Vujcic Z
(2008) Purification and properties of major midgut leucyl
aminopeptidase of Morimus funereus (Coleoptera, Cerambyci-
dae) larvae. Comp Biochem Physiol 149B:454–462
Bradford MM (1976) A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principle of protein-dye binding. Anal Biochem 72:248–254
Campanaro A, Bardiani M, Spada L, Carnevali L, Montalto F,
Antonini G, Audisio P, Mason F (2010) National guidelines for
monitoring and conservation of saproxylic coleopterans in Italy:
a contribution to implementing the habitats directive. Book of
abstracts, 6th European symposium on the conservation of
saproxylic beetles, Ljubljana, pp 14–15
Council of Europe (1996) Background information on invertebrates of
the habitats directive and the bern convention, nature and
environment, vol 79. Council of Europe Publishing, Strasbourg,
pp 59–62
Decree on designate and protect the strictly protected and endangered
species of wild plants, animals and fungi (2010) Sluzbeni glasnik
5/10 [in Serbian] pp 46
Dojnov B, Bozic N, Nenadovic V, Ivanovic J, Vujcic Z (2008)
Purification and properties of midgut a-amylase isolated from
Morimus funereus (Coleoptera: Cerambycidae) larvae. Comp
Biochem Physiol 149B:153–160
Ðordevic S, Nenadovic V, Ivanovic J (1999) Effect of crude corpora
cardiaca extracts on carbohydrate and lipid metabolism in larvae
of a cerambycid beetle Morimus funereus as a function of diet
and temperature. Comp Biochem Physiol 122A:191–198
Gullan PJ, Cranston PS (2005) The insects: an outline of entomology,
3th edn edn. Blackwell Publishing, Oxford
Hanks LM, McElfresh JS, Millar JG, Paine TD (1993) Phoracantha
semipunctata (Coleoptera: Cerambycidae), a serious pest of
Eucalyptus in California: biology and laboratory—rearing pro-
cedures. Entomol Soc Am 7:96–102
Ilic N (2005) Longhorn beetles (Coleoptera, Cerambycidae) from
Serbia. Faunistic review, SZGR«Joksimovic», Belgrade [in
Serbian] pp 15–16
Ilijin L, Jankovic-Tomanovic M, Mrdakovic M, Vlahovic M, Peric-
Mataruga V, Lazarevic J, Nenadovic V (2004) The influence of
dietary protein quality on midgut and brain proteins in Morimusfunereus larvae. Arch Biol Sci 56:9–13
IUCN (2009) Morimus funereus IUCN red list of threatened specie.
http://www.iucnredlist.org/apps/redlist/details/13875/0
Ivanovic J (1968) Basic characteristic of the succession of insect
populations in the process of destruction of oak (Quercuspedunculata). Zbornik za prirodne nauke 35:108–123
Ivanovic J (1970) Basis of the oligophagy of the species Morimusfunereus. Arch Biol Sc 20:53–57
Ivanovic J (1991) Metabolic response to stressors. In: Ivanovic J,
Jankovic-Hladni M (eds) Hormones and metabolism in insect
stress. CRC Press, Boca Raton, pp 27–69
Ivanovic J, Jankovic-Hladni M, Milanovic M (1975) Effect of
constant temperature on survival rate, neurosecrection and
246 J Insect Conserv (2012) 16:239–247
123

endocrine cells, and digestive enzymes in Morimus funereuslarvae (Cerambycidae: Coleoptera). Comp Biochem Physiol
50A:125–130
Ivanovic J, Jankovic-Hladni M, Stanic V, Nenadovic V, Frusic M
(1989) The role of neurosecretion and metabolism in develop-
ment of an oligophagous feeding habit in Morimus funereuslarvae (Col.: Cerambycidae). Comp Biochem Physiol
94A:167–171
Jankovic-Hladni M, Chen A, Ivanovic J, Ðordevic S, Stanic V, Peric
V, Frusic M (1992) Effects of diet and temperature on Morimusfunereus larval hemolymph cation concentracion. Arch Insect
Biochem Physiol 20:205–214
Keena MA (2005) Pourable artificial diet for rearing Anoplophoraglabripennis (Coleoptera: Cerambycidae) and methods to opti-
mize larval survival and synchronize development. Ann Entomol
Soc Am 98(4):536–547
Kim MK, Kim JS, Han JH, Kim YJ, Yoon C, Kim GH (2006) Mating
behavior of Pine Sawyer, Monochamus saltuarius Gebler
(Coleoptera: Cerambycidae). J Asia Pacific Entomol 9:275–280
Kimoto T, Duthie-Holt M (2004) Cerambyx cerdo Linnaeus—great
Capricorn beetle. In: Exotic forest Insect guidebook, her majesty
in right of Canada (Canadian Food Inspection Agency).
http://www.inspection.gc.ca. pp 30–32
Laemmli UK (1970) Cleavage of structural proteins during the
assembly of the head of bacteriofage T4. Nature 227:680–685
Lee CY, Lo KC (1998) Rearing of Anoplophora macularia (Thom-
son) (Coleoptera: Cerambycidae) on artificial diets. Appl
Entomol Zool 33:105–109
Lekovic S, Lazarevic J, Nenadovic V, Ivanovic J (2001) The effect of
heat stress on the activity of A1 and A2 neurosecretory neurons
of Morimus funereus (Coleoptera: Cerambycidae) larvae. Eur J
Entomol 98:13–18
Lewis OT, Thomas CD (2001) Adaptations to captivity in the
butterfly Pieris brassicae (L.) and the implications for ex situconservation. J Insect Conserv 5:55–63
Linit MJ (1985) Continuous laboratory culture of Monochamuscarolinensis (Coleoptera: Cerambycidae) with notes on larval
development. Ann Entomol Soc Amm 78:212–213
Loncar N, Bozic N, Nenadovic V, Ivanovic J, Vujcic Z (2009)
Characterization of trypsin-like enzymes from the midgut of
Morimus funereus (Coleopterae: Cerambycidae) larvae. Arch
Biol Sci 61:713–718
Loncar N, Bozic N, Nenadovic V, Ivanovic J, Vujcic Z (2010)
Purification and properties of trypsin-like enzyme from the
midgut of Morimus funereus (Coleoptera, Cerambycidae) larvae.
Arch Insect Bioch Physiol 74:146–232
Michaud JP, Grant AK (2005) The biology and behavior of the
longhorned beetle, Dectes texanus on sunflower and soybean.
J Insect Sci 5:25–29
Morimus funereus, Habitats directive article 17 reporting (2009)
Annex II species of the EU habitats and species directive,
European Environment Agency
Nation J (2008) Insect physiology and biochemistry, 2nd edn. CRC
Press Taylor & Franis Group, Boca Raton
Nenadovic V, Ivanovic J, Stanic V, Jankovic-Hladni M (1986) Some
characteristics of differentiation of the embryonic neuroendo-
crine sysitem of the Morimus Asper funereus Muls. (Coleopte-
rae: Cerambycidae). Acta Entomol Jug 22:39–45
Pearce-Kelly P, Jones R, Clarke D, Walter C, Atjin P, Cunningham
AA (1998) The captive rearing of threatened orthoptera: a
comparation of two species’ breeding programmes at the
Zoological Society of London. J Insect Conserv 2:201–210
Pil N, Stankovic M (2006) Cerambycidae (Coleoptera) of the
Zasavica special nature reserve (Serbia). Acta Entomol Serb
11:34–43
Rogers D, Lewthwaite S, Dentener P (2002) Rearing huhu beetle
larvae, Prionoplus reticularis (Coleoptera: Cerambycidae) on
artificial diet. New Zeal J Zool 29:303–310
Romero-Samper J, Bahillo P (1993) Some observations on the
biology and distribution of Morimus asper (Sulzer, 1776)
(Coleoptera, Cerambycidae) in the Iberian Peninsula. Boln Asoc
esp Ent 17:103–122
Sama G (1988) Coleoptera Cerambycidae, catalogo topografico e
sinonimico, Fauna d’Italia. Calderini. Bologna XXXVI: 216
Schultz CB, Dzurisin JD, Russell C (2009) Captive rearing of Puget
blue butterflies (Icaricia icarioides blackmorei) and implications
for conservation. J Insect Conserv 13:309–315
Shintani Y, Munyiri FN, Ishikawa Y (2003) Change in significance of
feeding during larval development in the yellow—spotted
longicorn beetle, Psacothea hilaris. J Insect Physiol 49:975–981
Simoneta J (1989) Contributo alla conoscenza dei cerambicidi della
Calabria (Coleoptera, Cerambycidae). Boll Ass Romana Ento-
mol 43:49–53
Stanic V, Ivanovic J, Jankovic-Hladni M, Nenadovic V, Marovic R
(1985) Feeding habits, behaviour, ovipositon and longevity of
the adult cerambycid beetle Morimus asper funereus muls. (Col.,
Cerambycidae) under laboratory conditions. Acta Entomol Jug
21:87–94
Terra WR, Ferreira C (1994) Insect digestive enzymes: properties,
compartmentalization and function. Comp Biochem Physiol
109B:1–62
Vrezec A, Ambrozic S, Kapla A (2010) Biology and ecology of
flightless cerambycid Morimus funereus (Muslant, 1862) as a
background for monitoring application: laboratory and large-
scale field study. Book of abstracts, 6th European symposium on
the conservation of saproxylic beetles, Ljubljana, p 20
Wagner W, Mohrlen F, Schnetter W (2002) Characterization of the
proteolytic enzymes in the midgut of the European cockchafer,
Melolontha melolontha (Coleoptera: Scarabaeidae). Insect Bio-
chem Mol Biol 32:803–814
Wang Q, Shi G, Song D, Rogers D, Davis L, Chen X (2002)
Development, survival, body weight, longevity and reproductive
potential of Oemena hirta (Coleoptera: Cerambycidae) under
different rearing conditions. J Econ Entomol 95:563–569
Yoon H, Mah Y (1999) Life cycle of the mulberry longicorn beetle,
Apriona germani hope on an artificial diet. J Asia-Pacific
Entomol 2:169–173
J Insect Conserv (2012) 16:239–247 247
123





























