Embed Size (px)
Citation preview

Plant Physiol. (1994) 104: 339-347
A Role for Cytokinins in De-Etiolation in Arabidopsis'
det Mutants Have an Altered Response to Cytokinins
Joanne Chory*, Dennis Reinecke', Sopheak Sim, Tracy Washburn, and Mark Brenner
Plant Biology Laboratory, The Salk lnstitute for Biological Studies, San Diego, California 921 86-5800 (J.C., S.S., T.W.); and Department of Horticultural Sciences,
University of Minnesota, St. Paul, Minnesota 55108 (D.R., M.B.)
When grown in the absence of light, Arabidopsis thaliana de- etiolated (det) mutants develop many of the characteristics of light- grown plants, including the development of leaves and chloro- plasts, the inhibition of hypocotyl growth elongation, and elevated expression levels of light-regulated genes. We show here that dark- grown wild-type seedlings exhibit similar phenotypic traits if any one of a variety of cytokinins are present in the growth medium. We further show that the striking phenotype of det mutants is unlikely to be caused by different levels of cytokinins in these mutants. l h e three major Arabidopsis cytokinins, zeatin, zeatin riboside, and isopentenyladenosine, accumulate to similar levels in wild-type seedlings grown in either the light or the dark. There is no consistently different pattern for the levels of these cytokinins in wild-type versus detl or det2 mutants. However, detl and det2 have an altered response to cytokinin in a detached leaf senescence assay and in tissue culture experiments. A model is proposed in which light and cytokinins act independently or sequentially through common signal transduction intermediates such as DE11 and DET2 to control the downstream light-regulated responses.
Light effects on plant growth and development can be observed at almost every stage of the life cycle, from germi- nation to floral induction. In particular, light causes profound changes in the morphology of the developing young dicoty- ledonous seedling. These changes include production of Chl, differentiation of leaves and cotyledons, inhibition of stem growth elongation, and the development of the photosyn- thetically competent chloroplast. In the few cases for which there are available data, the molecular basis-for light-regu- lated development involves the modulation of gene expres- sion (for reviews, see Gilmartin et al., 1990; Chory, 1993).
' Supported by grants from the National Science Foundation, the Competitive Grants Program of the U.S. Department of Agriculture, the Samuel Roberts Noble Foundation (J.C.), and The Minnesota Agricultura1 Experimental Station (M.B.). T.W. is a postdoctoral fel- low of the American Cancer Society. Some of these results were originally presented at XLV Symposium of the Society for'Experi- mental Biology held in Glasgow, Scotland, in August 1990, and parts have appeared in the proceedings from this meeting (Chory et al., 1991a).
Present address: Department of Plant Science, University of Alberta, Edmonton, Alberta TtG2P5, Canada.
* Corresponding author; fax 1-619-558-6379. 339
The molecular mechanisms by which light regulates de- velopment are largely unknown. Except for the red/far-red absorbing photoreceptor, phytochrome, little is known about the biochemistry of signal perception and transduction (Quail, 1991). A further complication exists in the plethora of literature documenting that light and hormones cause similar effects in developing plants. For instance, control of gennination has been shown to involve GAs, cytokinin, and ABA in various species (Moore, 1979; Jacobsen and Chandler, 1987). GAs, auxins,'and ethylene are each involved in the control of cell elongation and morphological responses re- quired for seedling emergence from the soil (Evans, 1985; Potts et al., 1985; Abeles et al., 1992; Kieber et al., 1993). Cytokinins promote cotyledon expansion, leaf development, and chloroplast differentiation (Miller, 1956; Stetler and Laetsch, 1965; Huff and Ross, 1975). During later vegetative growth, cytokinins and ethylene control the onset of leaf senescence (Leopold and Kawase, 1964; Gepstein and Thi- mann, 1981). Finally, the ratio of cytokinins to auxins is a primary determinant in the control of apical dominance (Moore, 1979). The overlapping roles of light and plant hormones in development raises the interesting question of whether light and hormones act independently to affect developmental responses or whether plant hormones are involved in the sequence of events initiated by physiologically active photoreceptors.
We have taken a genetic approach to the study of how light interacts with endogenous developmental programs in developing young Arabidopsis seedlings. Our studies have identified a phenotypic class of mutants that show many characteristics of light-grown plants even when grown in complete darkness. We have designated these mutants det (for de-etiolated) because they are de-etiolated in the dark, instead of having the usual etiolated seedling morphology. Recessive mutations in any one of four DET3 genes result in
Genetic nomenclature: Wild-type gene symbols are capitalized (e.g., DETI, gene product, DETI) and mutants are represented with lowercase symbols (detl), as established at the Third Intemational Arabidopsis Meeting held at Michigan State University in 1987.
Abbreviations: (diH)Z, dihydrozeatin; (diH)[SR]Z, dihydrozeatin riboside; GUS, 8-glucuronidase; iP, isopentenyladenine; [9R]iP, iso- pentenyladenosine; Z, zeatin; [9G]Z, zeatin-9-glucoside; [9R]Z, zeatin riboside.
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.

340 Chory et al. Plant Physiol. Vol. 104, 1994
dark-grown plants that grow similarly to light-grown wild- type seedlings (Chory et al., 1989, 1991b; Cabrera et al., 1993; H. Cabrera and J. Chory, unpublished data).
We have been studying homozygous mutant detl and det2 seedlings in the most detail. Dark-grown detl and det2 plants have short hypocotyls, expanded cotyledons, and developed leaves. Additionally, severa1 light-regulated genes, including cab (codes for the light-harvesting Chl proteins of PSII), rbcS (codes for the small subunit of Rubisco), and chs (codes for chalcone synthase) are expressed at high levels in the dark in these mutants (Chory et al., 1989, 1991b). When grown in the light, detl and det2 are small and have reduced apical dominance and fertility when compared with wild-type Ar- abidopsis (Chory and Peto, 1990; Chory et al., 1991b). These data imply that the DETl and DET2 gene products play a role in light-grown plants, as well as in dark-grown plants. Specifically, light-grown plants appear to require DETl for spatial repression of light-regulated genes, because cab, chs, and rbcS are ectopically expressed in detl mutants (Chory and Peto, 1990). In contrast, homozygous recessive det2 mutations affect photoperiodic responses in light-grown plants, including delayed timing of leaf and chloroplast se- nescence, delayed flowering, and reduced dark adaptation of cab gene expression (Chory et al., 1991b). Therefore, DET2 may play a negative role in the temporal elaboration of light responses during Arabidopsis development. Epistasis analyses between detl and det2 and detl or det2 with phytochrome- deficient mutants indicate that DETl and DET2 act on distinct pathways and are signal transduction elements that act downstream from phytochrome to affect developmental re- sponses (Chory, 1992).
The pleiotropic nature of the detl and det2 mutations suggested our original proposal that these genes may play a role in central developmental pathways that respond to the light environment (Chory et al., 1989). This notion was further reinforced by an experiment in which we put detl tissue into culture on synthetic medium containing only auxin. We observed that detl calli were somewhat green instead of achlorophyllous like the wild type, indicating that the undifferentiated calli contained chloroplasts. Because cy- tokinin is normally required for greening, this observation suggested that detl mutants had become cytokinin autono- mous. Here, we test the possibility that aberrant cytokinin physiology is related to the numerous phenotypes observed in detl and det2 seedlings. We show that many of the phenotypes of de t l mutants can be mimicked by the addition of cytokinins to the growth medium; however, detl and det2 seedlings do not simply contain elevated cytokinin levels. Responses of de t l , det2, and wild-type explants to various concentrations of cytokinins suggest that an altered response to cytokinins is partially responsible for the de-etiolated phenotypes observed in det mutants. A model is proposed in which light and cytokinins act in concert to regulate either the pool size or activity of DETl and DET2, which in turn affects the elaboration of the downstream light-regulated developmental responses.
MATERIALS AND METHODS
Plant Material and Growth Conditions
A11 strains of Arabidopsis used in this study were ecotype Columbia. The de t l and det2 mutants have been described
previously (Chory et al., 1989, 1991b). Seedlings were grown aseptically on synthetic medium supplemented with 2% SUC and cytoltinins as indicated. Seeds were cold treated over- night and then germinated in high photon fluence rate white light at 20°C under a mixture of fluorescent and incandescent lights at an intensity of 350 pE m-2 s-’. Dark-grown seeds were germinated for 24 h in the light before transfer to total darkness for 7 to 21 d. A green safelight filter was used during a11 dark manipulations.
In the experiments described in Figure 1, hypocotyl elon- gation was measured with a ruler after growth for 7 d in the dark. At least 100 seedlings were measured for each sample.
Gene Expression Studies
The construction of the transgenic lines containing cab3- GUS and chs-GUS was published elsewhere (Chory and Peto, 1990). GLJS assays with protein extracts from dark- and light- grown seedlings were camed out using the fluoronietric assay (Jefferson, 1987).
EM
Cotyledon tissue from 8-d-old dark-grown seedlings was used as the source of material. Tissues were processed as described by Chory et al. (1991b).
Cytokinin Extraction and Measurement
The method of cytokinin measurement was similar to that described by Schreiber (1990) with the following modifica- tions. Tissues were frozen with liquid N2, ground with mortar and pestle, and homogenized in a Polytron with 5 mL g-’ fresh weight of extraction buffer (methanol:H20:acetic acid [70:27:3] plus 10 mg L-’ of butylated hydroxytoluene plus 10% PVP g g-’ fresh weight). [“C]Z, [3H][9R]Z, and [3H][9R] iP were added to extracts for recovery estimation. After 24 h of equilibration, the extract was pelleted and the pellet was reextracted. The combined supematant was dried, and the residue was treated with acid phosphatase. The extract was applied to a 2-mL CI8 Megabond elute cartridge and washed with 2 volumes of 0.1 M acetic acid (pH 3.3) with triethanol- amine (solvent B) and eluted with 2 volumes of methanol. The sample was dried and chromatographed over a 2-mL DEAE column onto a 1-mL Affigel anti-[9R]Z, (diH)[9R]Z, and [9R]iP antibody column (washed sequentially with 10 mM Bis Tris, pH 6.5 [buffer A], buffer A plus 1 M NaC1, H20, and methanol). The methanol eluant was dried, resuspended in solvent B plus 100 ng of kinetin, injected onto a Cls HPLC (0.45- X 25-cm column at 1.5 mL min-’), and eluted with a gradient of solvent A (CH3CN) and solvent B (9:91,0-9 min; 17233, 9-40 min; and 100:0, 40-48 min). Under these con- ditions cytokinins had retention times (min) of 5.3 for [9G]Z, 7.6 for Z, 8.5 for [diH]Z, 14.1 for [9R]Z, 14.8 for (diH)[9R]Z, 17.0 for kinetin, 26.1 for iP, and 29.1 for [9R]iP. Endogenous cytokinins (base and nucleoside plus nucleotide) were iden- tified and quantified by diode array detection and correc- ted for recovery of radiolabeled cytokinins by scintillation counting.
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.
Phenocopy of Arabidopsis det Mutants 341
Detached Leaf Senescence Assays
Leaf senescence assays were performed as described by W. Su and S. Howell (personal communication). Plants were grown on synthetic medium plus agar for approximately 3 weeks (Chory et al., 1989). Whole leaves from wild type, de t l , and det2 were then detached with a scalpel and placed into Petri dishes containing severa1 layers of filter paper saturated with water or solutions containing various levels of BA in water. The Petri dishes were incubated in the dark for 7 d, taking care to keep the leaves moist. Specific Chl content was calculated as follows. Two leaves per sample were blotted dry, weighed, and placed in a 1.5-mL Eppendorf tube. The samples were resuspended in 80% acetone, ground with a disposable pestle, and incubated in the dark for 30 min. Total Chl (pg mL-') was determined according to the equation: 20.2 + 8.02 A663. At least 10 leaves per treatment were used.
Tissue Culture Experiments
Wild type, de t l , and der2 plants were grown aseptically on minimal salts medium plus Suc for 2 weeks (Chory et al., 1989). At that time, stem, root, and leaf explants were incu- bated for 4 d on callus induction medium (Valvekens et al., 1988). The explants were then transferred to various media with no hormones or different quantities of kinetin as de- scribed in "Results." Organogenesis was scored after 2 weeks of growth.
RESULTS
Exogenously Applied Cytokinins Result in a Wild-Type Phenocopy of detl Mutants
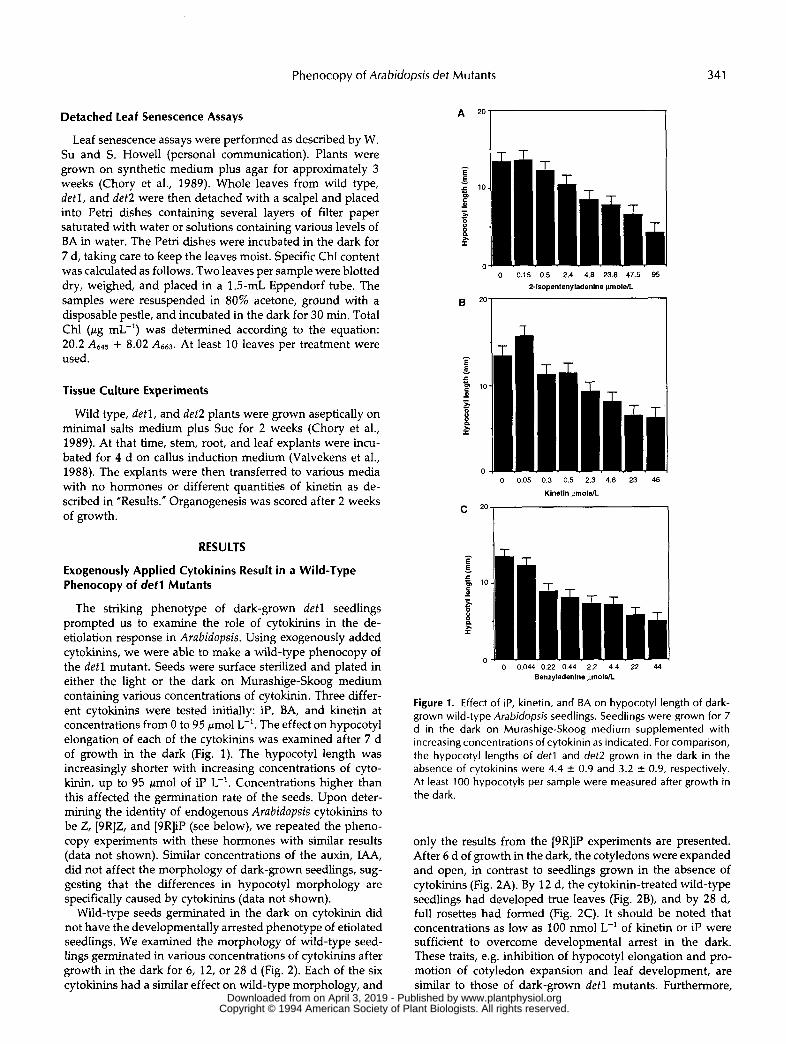
The striking phenotype of dark-grown detl seedlings prompted us to examine the role of cytokinins in the de- etiolation response in Arabidopsis. Using exogenously added cytokinins, we were able to make a wild-type phenocopy of the detl mutant. Seeds were surface sterilized and plated in either the light or the dark on Murashige-Skoog medium containing various concentrations of cytokinin. Three differ- ent cytokinins were tested initially: iP, BA, and kinetin at concentrations from O to 95 pmol L-'. The effect on hypocotyl elongation of each of the cytokinins was examined after 7 d of growth in the dark (Fig. 1). The hypocotyl length was increasingly shorter with increasing concentrations of cyto- kinin, up to 95 Imo1 of iP L-'. Concentrations higher than this affected the germination rate of the seeds. Upon deter- mining the identity of endogenous Arabidopsis cytokinins to be Z, [9R]Z, and [9R]iP (see below), we repeated the pheno- copy experiments with these hormones with similar results (data not shown). Similar concentrations of the auxin, IAA, did not affect the morphology of dark-grown seedlings, sug- gesting that the differences in hypocotyl morphology are specifically caused by cytokinins (data not shown).
Wild-type seeds germinated in the dark on cytokinin did not have the developmentally arrested phenotype of etiolated seedlings. We examined the morphology of wild-type seed- lings germinated in various concentrations of cytokinins after growth in the dark for 6, 12, or 28 d (Fig. 2) . Each of the six cytokinins had a similar effect on wild-type morphology, and
O 0.15 0.5 2.4 4.8 23.8 47.5 95 Blsopentenyladenlne &moleR
E 2o
T
O 0.05 0.3 0.5 2.3 4.6 23 46
Kineiln f i m o l d
I,
O 0044 022 044 2 2 4 4 22 44 Benzyladenine pmolelL
Figure 1. Effect of iP, kinetin, and BA on hypocotyl length of dark- grown wild-type Arabidopsis seedlings. Seedlings were grown for 7 d in the dark on Murashige-Skoog medium supplemented with increasing concentrations of cytokinin as indicated. For comparison, the hypocotyl lengths of detl and det2 grown in the dark in the absence of cytokinins were 4.4 & 0.9 and 3.2 + 0.9, respectively. At least 100 hypocotyls per sample were measured after growth in the dark.
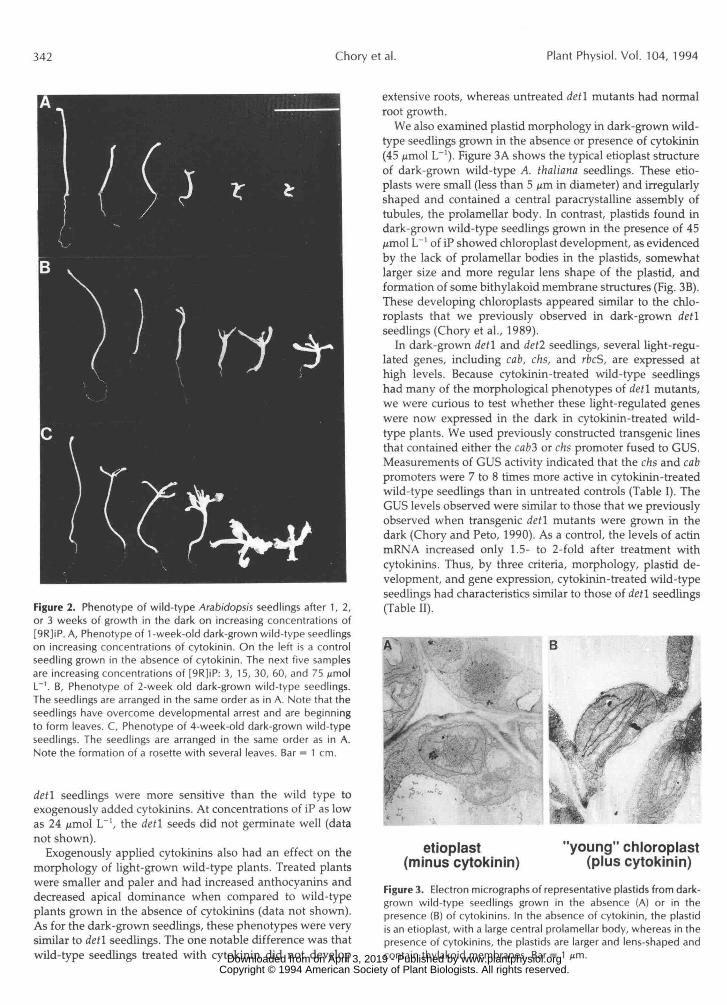
only the results from the [9R]iP experiments are presented. After 6 d of growth in the dark, the cotyledons were expanded and open, in contrast to seedlings grown in the absence of cytokinins (Fig. 2A). By 12 d, the cytokinin-treated wild-type seedlings had developed true leaves (Fig. 2B), and by 28 d, full rosettes had formed (Fig. 2C). It should be noted that concentrations as low as 100 nmol L-' of kinetin or iP were sufficient to overcome developmental arrest in the dark. These traits, e.g. inhibition of hypocotyl elongation and pro- motion of cotyledon expansion and leaf development, are similar to those of dark-grown det l mutants. Furthermore,
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.
342 Chory et al. Plant Physiol. Vol. 104, 1994
Figure 2. Phenotype of wild-type Arabidopsis seedlings after 1, 2,or 3 weeks of growth in the dark on increasing concentrations of[9R]iP. A, Phenotype of 1 -week-old dark-grown wild-type seedlingson increasing concentrations of cytokinin. On the left is a controlseedling grown in the absence of cytokinin. The next five samplesare increasing concentrations of [9R]iP: 3, 15, 30, 60, and 75 ^molL"1. B, Phenotype of 2-week old dark-grown wild-type seedlings.The seedlings are arranged in the same order as in A. Note that theseedlings have overcome developmental arrest and are beginningto form leaves. C, Phenotype of 4-week-old dark-grown wild-typeseedlings. The seedlings are arranged in the same order as in A.Note the formation of a rosette with several leaves. Bar = 1 cm.
detl seedlings were more sensitive than the wild type toexogenously added cytokinins. At concentrations of iP as lowas 24 ftmol L"1, the detl seeds did not germinate well (datanot shown).
Exogenously applied cytokinins also had an effect on themorphology of light-grown wild-type plants. Treated plantswere smaller and paler and had increased anthocyanins anddecreased apical dominance when compared to wild-typeplants grown in the absence of cytokinins (data not shown).As for the dark-grown seedlings, these phenotypes were verysimilar to detl seedlings. The one notable difference was thatwild-type seedlings treated with cytokinin did not develop
extensive roots, whereas untreated detl mutants had normalroot growth.
We also examined plastid morphology in dark-grown wild-type seedlings grown in the absence or presence of cytokinin(45 fimol L"1). Figure 3A shows the typical etioplast structureof dark-grown wild-type A. thaliana seedlings. These etio-plasts were small (less than 5 /*m in diameter) and irregularlyshaped and contained a central paracrystalline assembly oftubules, the prolamellar body. In contrast, plastids found indark-grown wild-type seedlings grown in the presence of 45jrniol L"1 of iP showed chloroplast development, as evidencedby the lack of prolamellar bodies in the plastids, somewhatlarger size and more regular lens shape of the plastid, andformation of some bithylakoid membrane structures (Fig. 3B).These developing chloroplasts appeared similar to the chlo-roplasts that we previously observed in dark-grown detlseedlings (Chory et al., 1989).
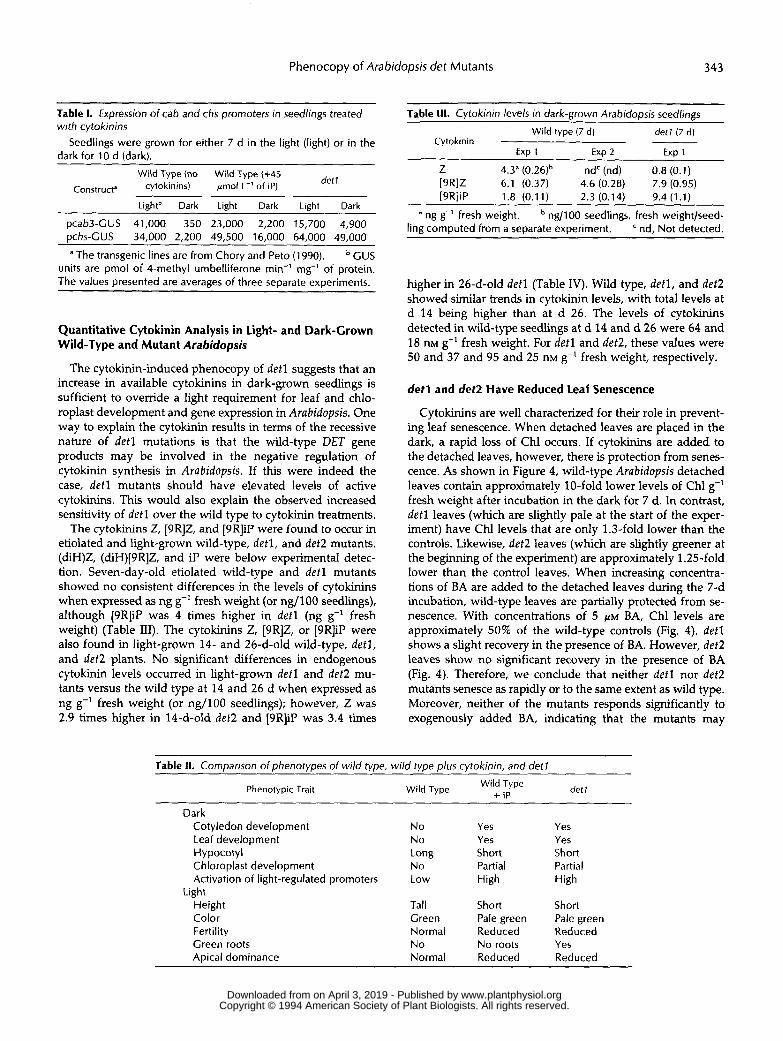
In dark-grown detl and det2 seedlings, several light-regu-lated genes, including cab, chs, and rbcS, are expressed athigh levels. Because cytokinin-rreated wild-type seedlingshad many of the morphological phenotypes of detl mutants,we were curious to test whether these light-regulated geneswere now expressed in the dark in cytokinin-rreated wild-type plants. We used previously constructed transgenic linesthat contained either the cab3 or chs promoter fused to GUS.Measurements of GUS activity indicated that the chs and cabpromoters were 7 to 8 times more active in cytokinin-treatedwild-type seedlings than in untreated controls (Table I). TheGUS levels observed were similar to those that we previouslyobserved when transgenic detl mutants were grown in thedark (Chory and Peto, 1990). As a control, the levels of acrinmRNA increased only 1.5- to 2-fold after treatment withcytokinins. Thus, by three criteria, morphology, plastid de-velopment, and gene expression, cytokinin-treated wild-typeseedlings had characteristics similar to those of detl seedlings(Table II).
B
etioplast(minus cytokinin)
'young" chloroplast(plus cytokinin)
Figure 3. Electron micrographs of representative plastids from dark-grown wild-type seedlings grown in the absence (A) or in thepresence (B) of cytokinins. In the absence of cytokinin, the plastidis an etioplast, with a large central prolamellar body, whereas in thepresence of cytokinins, the plastids are larger and lens-shaped andcontain thylakoid membranes. Bar = 1 ̂ m. www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from
Copyright © 1994 American Society of Plant Biologists. All rights reserved.
Phenocopy of Arabidopsis der Mutants 343
~ ~~~
Table I. Expressron of cab and chs promoters rn seedlrngs treated wrth cytok,nms
Seedlings were grown for either 7 d in the light (Iight) or in the dark for 10 d (dark).
detl Wild Type (no Wild Type (+45 Constructa cytokinins) pmol L-' of IP)
Lightb Dark Light Dark Light Dark
pcab3-CUS 41,000 350 23,000 2,200 15,700 4,900 pchs-CUS 34,000 2,200 49,500 16,000 64,000 49,000
a The transgenic lines are from Chory and Peto (1990) CUS units are pmol of 4-methyl umbelliferone min-' mg-' of protein. The values presented are averages of three separate experiments.
Quantitative Cytokinin Analysis in Light- and Dark-Crown Wild-Type and Mutant Arabidopsis
The cytokinin-induced phenocopy of der1 suggests that an increase in available cytokinins in dark-grown seedlings is sufficient to override a light requirement for leaf and chlo- roplast development and gene expression in Arabidopsis. One way to explain the cytokinin results in terms of the recessive nature of detl mutations is that the wild-type DET gene products may be involved in the negative regulation of cytokinin synthesis in Arabidopsis. If this were indeed the case, det l mutants should have elevated levels of active cytokinins. This would also explain the observed increased sensitivity of det 1 over the wild type to cytokinin treatments.
The cytokinins Z, [9R]Z, and [9R]iP were found to occur in etiolated and light-grown wild-type, detl, and det2 mutants. (diH)Z, (diH)[BR]Z, and iP were below experimental detec- tion. Seven-day-old etiolated wild-type and detl mutants showed no consistent differences in the levels of cytokinins when expressed as ng g-' fresh weight (or ng/100 seedlings), although [9R]iP was 4 times higher in detl (ng g-' fresh weight) (Table 111). The cytokinins Z, [9R]Z, or [9R]iP were also found in light-grown 14- and 26-d-old wild-type, detl, and det2 plants. No significant differences in endogenous cytokinin levels occurred in light-grown detl and det2 mu- tants versus the wild type at 14 and 26 d when expressed as ng g-' fresh weight (or ng/100 seedlings); however, Z was 2.9 times higher in 14-d-old det2 and [9R]iP was 3.4 times
Table 111. Cytokinin levels in dark-grown Arabidopsis seedlings Wild type (7 d) detr (7 d)
Cytokinin Exp 1 Exp 2 Exp 1
Z 4.3" (0.26)b n d ' (nd) 0.8 (0.1) [9RlZ 6.1 (0.37) 4.6 (0.28) 7.9 (0.95) [9R]iP 1.8 (0.11) 2.3 (0.14) 9.4 (1.1)
a ng g-' fresh weight. ng/100 seedlings, fresh weightlseed- nd, Not detected. ling computed from a separate experiment.
higher in 26-d-old detl (Table IV). Wild type, detl, and det2 showed similar trends in cytokinin levels, with total levels at d 14 being higher than at d 26. The levels of cytokinins detected in wild-type seedlings at d 14 and d 26 were 64 and 18 nM g-' fresh weight. For detl and det2, these values were 50 and 37 and 95 and 25 nM g-' fresh weight, respectively.
detl and det2 Have Reduced Leaf Senescence
Cytokinins are well characterized for their role in prevent- ing leaf senescence. When detached leaves are placed in the dark, a rapid loss of Chl occurs. If cytokinins are added to the detached leaves, however, there is protection from senes- cence. As shown in Figure 4, wild-type Arabidopsis detached leaves contain approximately 10-fold lower levels of Chl g-' fresh weight after incubation in the dark for 7 d. In contrast, detl leaves (which are slightly pale at the start of the exper- iment) have Chl levels that are only 1.3-fold lower than the controls. Likewise, det2 leaves (which are slightly greener at the beginning of the experiment) are approximately 1.25-fold lower than the control leaves. When increasing concentra- tions of BA are added to the detached leaves during the 7-d incubation, wild-type leaves are partially protected from se- nescence. With concentrations of 5 PM BA, Chl levels are approximately 50% of the wild-type controls (Fig. 4). detl shows a slight recovery in the presence of BA. However, det2 leaves show no significant recovery in the presence of BA (Fig. 4). Therefore, we conclude that neither detl nor det2 mutants senesce as rapidly or to the same extent as wild type. Moreover, neither of the mutants responds significantly to exogenously added BA, indicating that the mutants may
Table II. Comparison ofphenotypes of wild type, wild type plus cytokinin, and detl
Wild Type Wild Type det 1 + iP Phenotypic Trait
Dark Cotyledon development N o Ye5 Yes Leaf development No Yes Yes H ypocotyl Long Short Short Chloroplast development No Partial Partial Activation of light-regulated promoters Low High High
Height Tal1 Short Short Color Green Pale green Pale green Fertility Normal Reduced Reduced Green roots N o N o roots Yes Apical dominance Normal Reduced Reduced
Light
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.
344 Chory et al. Plant Physiol. Vol. 104, 1994
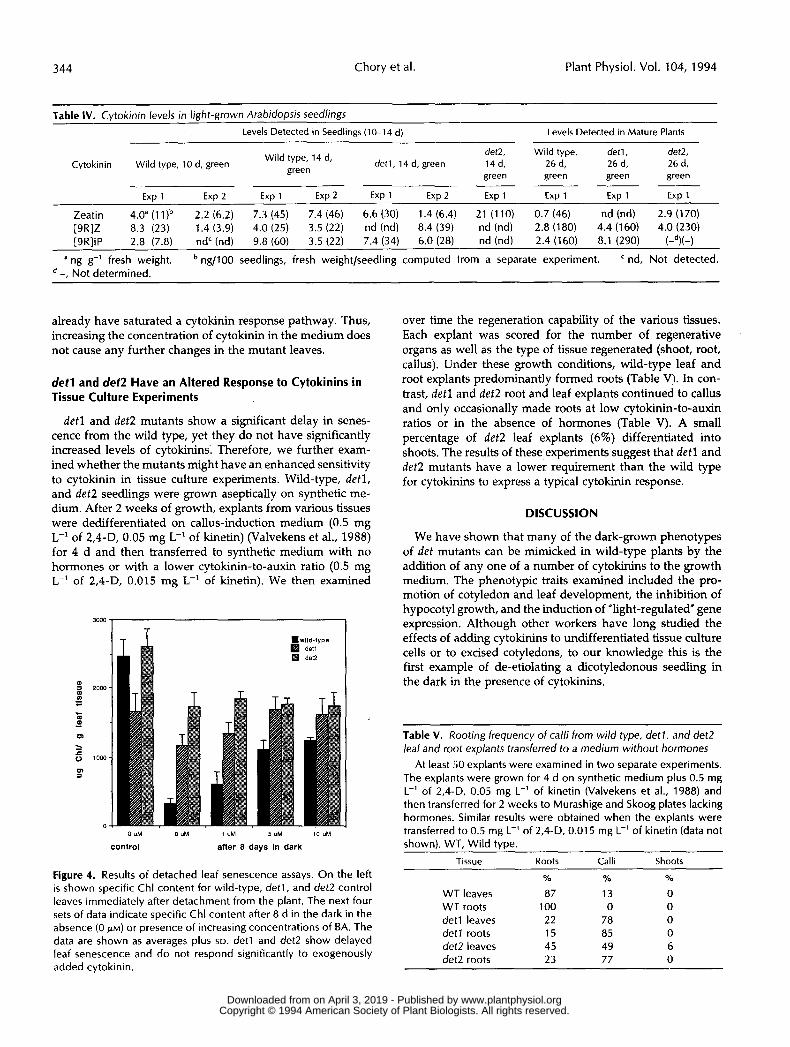
Table IV. Cytokinin levels in light-grown Arabidopsis seedlings Levels Detected in Seedlings (10-14 d) Levels Detected in Mature Plarits
det2, Wild type, detl, det2, Cytokinin Wild type, 10 d, green I._,,.. detl, 14 d, green 14 d, 26 d, 26 d, 26 d, Wild type, 14 d,
green green green green
Exp 1 Exp 2 Exp 1 Exp 2 Exp 1 Exp 2 Exp 1 Exp 1 Exp 1 Exp 1
5, CCI I
Zeatin 4.0" (1 l )b 2.2 (6.2) 7.3 (45) 7.4 (46) 6.6 (30) 1.4 (6.4) 21 (1 10) 0.7 (46) nd (nd) 2.9 (170) [9R]Z 8.3 (23) 1.4 (3.9) 4.0 (25) 3.5 (22) nd (nd) 8.4 (39) nd (nd) 2.8 (180) 4.4 (160) 4.0 (230) [9R]iP 2.8 (7.8) nd' (nd) 9.8 (60) 3.5 (22) 7.4 (34) 6.0 (28) nd (nd) 2.4 (160) 8.1 (290) (-d)(-)
a ng g-' fresh weight. ng/100 seedlings, fresh weightlseedling computed írom a separate experiment. nd, Not detected. -, Not determined.
already have saturated a cytokinin response pathway. Thus, increasing the concentration of cytokinin 'in the medium does not cause any further changes in the mutant leaves.
detl and det2 Have an Altered Response to Cytokinins in Tissue Culture Experiments
detl and det2 mutants show a significant delay in senes- cence from the wild type, yet they do not have significantly increased levels of cytokinins. Therefore, we further exam- ined whether the mutants might have an enhanced sensitivity to cytokinin in tissue culture experiments. Wild-type, detl, and det2 seedlings were grown aseptically on synthetic me- dium. After 2 weeks of growth, explants from various tissues were dedifferentiated on callus-induction medium (0.5 mg L-' of 2,4-D, 0.05 mg L-' of kinetin) (Valvekens et al., 1988) for 4 d and then transferred to synthetic medium with no hormones or with a lower cytokinin-to-auxin ratio (0.5 mg L-' of 2,4-D, 0.015 mg L-' of kinetin). We then examined
30W
@
v)
a 2000
- - I
al - a
" 1000
m
1 c
O
T mwi ld - t ype deí1
O UM O UM 1 UM 5 UM 10 UM
ontrol after 8 days in dark
over time the regeneration capability of the various tissues. Each explant was scored for the number of regenerative organs as well as the type of tissue regenerated (shoot, root, callus). Under these growth conditions, wild-type leaf and root explants predominantly formed roots (Table V). In con- trast, detl and det2 root and leaf explants continued to callus and only occasionally made roots at low cytokinin-to-auxin ratios or in the absence of hormones (Table V). A small percentage of det2 leaf explants (6%) differentiated into shoots. The results of these experiments suggest that detl and det2 mutants have a lower requirement than the wild type for cytokinins to express a typical cytokinin response.
DISCUSSION
We have shown that many of the dark-grown phenotypes of det mutants can be mimicked in wild-type plants by the addition of any one of a number of cytokinins to the growth medium. The phenotypic traits examined included the pro- motion of cotyledon and leaf development, the inhibition of hypocotyl growth, and the induction of 'light-regulated" gene expression. Although other workers have long studied the effects of adding cytokinins to undifferentiated tissue culture cells or to excised cotyledons, to our knowledge this is the first example of de-etiolating a dicotyledonous seedling in the dark in the presence of cytokinins.
Table V. Rooting frequency of calli from wild type, detl, and det2 leaf and root explants transferred to a medium without hormones
At least 50 explants were examined in two separate experiments. The explants were grown for 4 d on synthetic medium plus 0.5 mg L-' of 2,4-D, 0.05 mg L-' of kinetin (Valvekens et al., 1988) and then transferred for 2 weeks to Murashige and Skoog plates lacking hormones. Similar results were obtained when the explants were transferred to 0.5 mg L-' of 2,4-D, 0.01 5 mg L-' of kinetin (data not shown). WT, Wild type.
Tissue Roots Calli Shoots Figure 4. Results of detached leaf senescence assays. On the left is shown specific Chl content for wild-type, detl, and det2 control leaves immediately after detachment from t h e plant. The next four sets of data indicate specific Chl content after 8 d in the dark in t h e absence (O PM) or presence of increasing concentrations of BA. The data are shown as averages plus SD. detl and det2 show delayed leaf senescence and do not respond significantly to exogenously added cytokinin.
~ ~
% % %
WT leaves 87 13 O WT roots 1 O0 O O detl leaves 22 78 O detl roots 15 85 O det2 leaves 45 49 6 det2 roots 23 77 o
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.

Phenocopy of Arabidopsis det Mutants 345
Severa1 laboratories have previously shown that cytokinins increase the accumulation of proteins encoded by cab and rbcS (Fierabend and deBoer, 1978; Teyssendier de la Serve et al., 1985). More recently, Lemna gibba and cucumber have also been used to study cytokinin effects on light-regulated gene expression (Flores and Tobin, 1986, 1988; Cotton et al., 1990). From the Lemna studies, it was concluded that kinetin regulation of cab and rbcS mRNA levels was primarily post- transcriptional. In cucumber, the leve1 of phyA mRNA, a gene whose expression is negatively regulated by light, was deter- mined in etiolated seedlings treated with BA. These authors showed that addition of exogenous BA to etiolated seedlings resulted in a significant decrease in phyA mRNA accumula- tion. This decrease was of the same order of magnitude as that caused by a red-light pulse, suggesting that BA can completely substitute for phytochrome in down-regulating the accumulation of phyA mRNA (Cotton et al., 1990). In Arabidopsis, the increased accumulation of cab and chs must be at least partially due to increases in transcription rates, because we have shown that the promoter is a11 that is required for increased activity of either the chloramphenicol acetyl transferase (data not shown) or the GUS marker genes.
The cytokinin phenocopy results imply that an increase in available cytokinins in dark-grown seedlings is sufficient to ovemde a light requirement for leaf and chloroplast devel- opment and gene expression in Arabidopsis. There are several ways to explain the cytokinin results in terms of the recessive nature of det mutations. First, DETl and DET2 may play a negative role in a cytokinin signal transduction pathway that is coupled to photoperception. A second altemative is that DETl and DETZ conjugate cytokinins, making them inacces- sible, or that they modify active cytokinins, creating an inactive form (e+ cytokinin glucosides). Third, DETl and DETZ may be involved in the negative regulation of cytokinin synthesis in Arabidopsis.
Quantitative cytokinin analysis in Arubidopsis seedlings indicates that Arabidopsis contains relatively low levels of the cytokinins Z, [9R]Z, and [9R]iP. If detl was a cytokinin biosynthesis overproduction mutant, the levels of cytokinins in dark-grown det l would be expected to be consistently higher than in wild-type etiolated seedlings. For instance, in the moss, Physcomitrellu patens, cytokinin levels are up to 100-fold higher than wild type in overproducing mutant lines (Wang et al., 1981a, 1981b). However, cytokinin levels did not show a consistent difference between dark-grown detl and wild-type Arabidopsis. Light-grown wild-type, detl, and det2 mutants also had no consistent difference in cytokinin levels. Although we did make several measurements in light- grown plants, we did not measure kinetics of accumulation of cytokinins throughout the entire developmental time or immediately after transfer of seedlings from dark to light growth conditions. There are reports that cytokinin levels increase transiently after red-light pulses in Scots pine seeds and in etiolated squash seedlings (Qamaruddm and Tillberg, 1989; Kuraishi et al., 1991). Therefore, it is possible that there could be a significant difference in cytokinin levels in the det mutants versus the wild type if measurements were made during the proper "developmental window."
A second caveat is that the cytokinin levels measured are averages for the entire plant. Because the phenotype of det
mutants is expressed in many organs of the mature plant, including roots, leaves, and stems, an increase in cytokinin levels might be expected to occur in several tissues simulta- neously . Our experiments do not eliminate the possibility that cytokinins are redistributed in response to light, even though the absolute levels remain the same. The phenotype of dark-grown det mutants might be explained by redistri- bution of a cytokinin in the absence of light signals. However, complete cytokinin measurements during development and among various tissues would be extremely difficult given the small size of Arabidopsis seedlings and the low endogenous levels of cytokinins. Indeed, tens of thousands of mutant plants were needed to perform the cytokinin quantitation presented here.
We believe that the phenocopy results are significant, even though extremely high quantities of iP were required to see the most dramatic phenotypic differences. Our most striking phenocopy results occurred in the range of 20 to 50 pmol L-' of iP, which is about 1000-fold higher than the endogenous levels. However, iP is not a very active cytokinin, and release from developmental arrest occurred at concentrations of iP as low as 100 nmol L-'. Moreover, we do not know the amount of exogenously supplied cytokinin actually taken up into dark-grown wild-type seedlings.
Campbell and Town (1991) measured cytokinin levels in wild-type and radiation-induced tumors of Arabidopsis thal- iana, ecotype Landsberg, using immunoaffinity columns and HPLC. The 8-d-old, light-grown wild type was observed to have [9R]Z and [9R]iP at 0.5 to 4.2 and 1.5 to 2.9 ng g-' fresh weight, respectively, levels similar to 1 O-d-old, light-grown wild-type Columbia seedlings used in our studies. The au- thors concluded that the y-radiation-induced tumors (includ- ing a shooty tumor) had cytokinin levels similar to control tissues. Conversely, crown gall shooty tumors had 100 to 1000 times higher levels of cytokinins than wild-type tissues. Clearly, det mutants are not simply cytokinin overproducers as are the Agrobacterium-transformed Arabidopsis shooty tu- mors of Campbell and Town. The possible biological signifi- cance of the relatively small cytokinin changes observed in det mutants is unclear.
Recent studies involving recombinant Agrobacterium rhi- zogenes rolC protein suggest that the rolC gene product transforms plants by releasing cytokinins from inactive N- glucoside conjugates (Estruch et al., 1991). By corollary, re- sults of the present study infer the presence of an activity or activities that modifies active cytokinins such that they are inaccessible to the plant. In our studies we examined tissue for the presence of cytokinin N-glucosides; however, signifi- cant levels of N-glucosides were not observed. Cytokinin O- glucosides were not measured in this study. The current quantitative cytokinin analysis provides no evidence for the hypothesis that DETl or DET2 modifies active cytokinins into inactive forms. Thus, a regulatory role for DETl or DETZ as an "anti-rolC" activity is not likely.
We favor a model in which light and cytokinins act inde- pendently or sequentially through common signal transduc- tion intermediates, such as DETl and DET2 to control the downstream light-regulated responses (Fig. 5, A and B). The primary mode of action of phytochrome, the blue-light or UV photoreceptors, or cytokinins is currently unknown.
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.

346 Chory et al. Plant Physiol. Vol. 104, 1994
A Light LI DET, 'I / Leaf deveiopment
Chioroolast differentiation
Cytokinins \ Gene expression
A Leaf development
Light--) Cytokinins 5 Chioropiast dilferentiation \ DETZ \ Gene expression
Leaf deveiopment
7 :::i} c$z:F /d Chloroplast dilferentiation
\ Gene exDression
Figure 5. Models to explain the det phenocopy results. A, DETl and DETZ are negatively acting components on a signal transduction pathway that is modulated independently by both light and cyto- kinins. B, Light is postulated to act through cytokinins to down- regulate the activities or concentrations of DETl and DETZ. C, DETl and DETZ act to link photoreceptors to a cytokinin signal transduc- tion pathway that regulates t h e downstream light-regulated responses.
Based on their data with red-light and cytokinin treatments, Flores and Tobin (1986) proposed that phytochrome and cytokinins independently change the pool size of a common intermediate, and this intermediate more directly regulates gene expression. These intermediates could be specified by the wild-type DETl and DET2 genes. Our limited data are consistent with the interpretation that light may act through DETl and DETZ to induce enhanced sensitivity to existing endogenous cytokinin levels (Trewavas, 1983). Equally plau- sible is a model in which light or cytokinins act to decrease the activity of DETl or DET2, which then permits the elab- oration of the downstream light-regulated gene expression and morphological responses. Finally, it is possible that DETl or DETZ act to link photoreceptors to a cytokinin action pathway (Fig. 5C).
The question of whether light and cytokinins act inde- pendently to affect developmental responses or whether plant hormones are involved in the sequence of events initi- ated by physiologically active photoreceptors is still an open one. For instance, in the Lemna studies, exogenously added cytokinin resulted in increased accumulation of cab and rbcS mRNAs to approximately the same levels as when short red- light treatments were applied. However, it was concluded that red light does not act by a cytokinin-mediated mecha- nism on cab and rbcS expression, because red light or cyto- kinin was each more effective in the presence of the other than when administered alone (Flores and Tobin, 1986). This result is in agreement with studies designed to measure increases in Chl accumulation in greening mustard cotyledons in response to red light and cytokinins (Mohr, 1984). Mohr and co-workers found a simple additive interaction when cytokinins and light were applied simultaneously and con- cluded that the two factors act on different causal events
leading to Chl accumulation. In contrast, in cucumber, phyA mRNA levels are down-regulated to similar levels by either light or BA (Cotton et al., 1990). Our studies do not directly address this question; however, the observation that cytoki- nin can completely override the necessity for light to induce leaf and chloroplast development and gene expression sug- gests that these responses are not additive in Arabidopsis.
High cytokinin-to-auxin ratios are known to promote greening and shoot formation from callus tissue in many plants. detl calli appear to be more sensitive to low ratios of cytokinins to auxin, because they have increased specific Chl content over wild type on low concentrations of cytokinin (data not shown). Even more striking was the laclc of root formation in detl and det2 seedlings transferred to a minimal salts medium after callus induction. This result implies that calli derived from detl and det2 explants have a lower re- quirement for cytokinins to express typical cytolunin re- sponses. Alternatively, the mutants may have lower cytokinin catabolism, and the cytokinins are more persistent after trans- fer of the explants to medium containing no hormones. Thus, det mutants may be opposite to recently isolated cytokinin- resistant mutants (ckr) that have a reduced response to cyto- kinins (Su and Howell, 1992). In contrast to det2 mutants, the ckr mutants are pale and have longer than normal primary roots. Moreover, auxin-resistant mutants of Arubidopsis (axr) have been described that have decreased responses to auxin (Estelle and Somerville, 1987). When double mutants were made between detl and uxrl, or det2 and nxrl, we observed a much more severe phenotype than for either mutant alone (J. Reed and J. Chory, unpublished data). It will be informa- tive to make the detl-ckr and det2-ckr double mutants as well.
ACKNOWLEDCMENTS
We thank Marc Lieberman for photographing the seedlings in Figure 2, Charles Peto and AbbyAnn Sisk for the electron micro- graphs shown in Figure 3, and Cynthia Doane for help in assembling this manuscript. Dr. Mich Hein (Scripps Research Institute) provided valuable advice during the early stages of this work.
Received July 1, 1993; accepted September 15, 1993. Copyright Clearance Center: 0032-0889/94/104/0339/09.
LITERATURE CITED
Abeles FB, Morgan PW, Saltveit ME (1992) Ethylene in Plant Biology, Ed 2. Academic Press, San Diego, CA
Cabrera H, Peto C, Chory J (1993) A mutation in the Arabidopsis DET3 gene uncouples photoregulated leaf development from gene expression and chloroplast biogenesis. Plant J 4 671-682
Campbell BR, Town CD (1991) Physiology of hormone autonomous tissue lines derived from radiation-induced tumors of Arabidopsis fhaliana. Plant Physiol97: 1166-1 173
Chory J (1992) A genetic model for light-regulated seedling devel- opment in Arabidopsis. Development 115 337-354
Chory J (1993) Out of darkness: mutants reveal pathways controlling light-regulated development in plants. Trends Genet 9: 167-1 72
Chory J, Aguilar N, Peto C (1991a) The phenotype of Arabidopsis thaliana det l mutants suggests a role for cytokinins in greening. Zn GI Jenkins, W Schuch, eds, Molecular Biology of Plant Develop- ment, Symposia of the Society for Experimental Biology, No XLV. The Company of Biologists Ltd, Cambridge, UK, pp 21-29
Chory J, Nagpal P, Peto C (1991b) Phenotypic and genetic analysis
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.

Phenocopy of Arabidopsis det Mutants 347
of det2, a new mutant that affects light-regulated seedling devel- opment in Arabidopsis. Plant Cell 3 445-459
Chory J, Peto C (1990) Mutations in the DETl gene affect cell-type- specific expression of light-regulated genes and chloroplast devel- opment in Arabidopsis. Proc Natl Acad Sci USA 87: 8776-8780
Chory J, Peto C, Feinbaum R, Pratt L, Ausubel F (1989) Arabidopsis thaliana mutant that develops as a light-grown plant in the absence of light. Cell58 991-999
Cotton J, Ross C, Byrne D, Colbert J (1990) Down-regulation of phytochrome mRNA abundance by red light and benzyladenine in etiolated cucumber cotyledons Plant Mo1 Biol 1 4 707-714
Estelle M, Somerville C (1987) Auxin-resistant mutants of Arabi- dopsis thaliana with an altered morphology. Mo1 Gen Genet 206
Estruch JJ, Chriqui D, Grosmann K, Schell J, Spena A (1991) The plant oncogene rolC is responsible for the release of cytokinins from glucoside conjugates. EMBO J 10: 2889-2895
Evans ML (1985) The action of auxin on plant cell elongation. Crit Rev Plant Sci 2 213-265
Fierabend J, deBoer J (1978) Comparative analysis of the action of cytokinin and light in the formation of ribulosebisphosphate car- boxylase and plastid biogenesis. Planta 142 75-82
Flores S, Tobin E (1986) Benzyladenine modulation of the expression of two genes for nuclear-encoded chloroplast proteins in Lemna gibba: apparent post-transcriptional regulation. Planta 168:
Flores S, Tobin E (1988) Cytokinin modulation of LHCP mRNA levels: the involvement of post-transcriptional regulation. Plant Mo1 Biol 11: 409-415
Gepstein S, Thimann KV (1981) The role of ethylene in the senes- cence of oat leaves. Plant Physiol 6 8 349-354
Gilmartin P, Sarokin L, Memelink J, Chua N-H (1990) Molecular light switches for plant genes. Plant Cell 2 369-378
Huff AK, Ross CW (1975) Promotion of radish cotyledon enlarge- ment and reducing sugar content by zeatin and red light. Plant Physiol56 429-433
Jacobsen JV, Chandler PM (1987) Gibberellin and abscisic acid in germinating seedlings. In PJ Davies, ed, Plant Hormones and Their Role in Plant Growth and Development. Kluwer, Boston, pp
Jefferson R (1987) Assaying chimeric genes in plants: the GUS gene fusion system. Plant Mo1 Biol Rep 5 387-405
Kieber J, Rothenberg M, Roman G, Feldmann K, Ecker J (1993) CTR1, a negative regulator of the ethylene response pathway in
200-206
340-349
164-193
Arabidopsis, encodes a member of the raf family of protein kinases. Cell72 427-441
Kuraishi S, Tasaki K, Sakurai N, Sadatoku K (1991) Changes in levels of cytokinins in etiolated squash seedlings after illumination. Plant Cell Physiol32 585-591
Leopold AC, Kawase M (1964) Benzyladenine effects on bean leaf growth and senescence. Am J Bot 51: 294-298
Miller C (1956) Similarity of some kinetin and red light effects. Plant Physiol31: 318-319
Mohr H (1984) Phytochrome and chloroplast development. ln NR Baker, J Barber, eds, Chloroplast Biogenesis. Elsevier, Amsterdam,
Moore TC (1979) Biochemistry and Physiology of Plant Hormones. Springer-Verlag, New York
Potts WC, Reid JB, Murfet IC (1985) Intemode length in Pisum. Gibberellins and the slender phenotype. Physiol Plant 63:
Qamaruddin M, Tillberg E (1989) Rapid effects of red light on the isopentenyladenosime content in Scots pine seeds. Plant Physiol
Quail P (1991) Phytochrome: a light-activated molecular switch that regulates plant gene expression. Annu Rev Genet B25: B389-B409
Schreiber BMN (1990) The role of cytokinins in maize kemel de- velopment. PhD dissertation. University of Minnesota, St Paul, MN
Stetler D, Laetsch W (1965) Kinetin induced chloroplast maturation in cultures of tobacco tissue. Science 149 1387-1389
Su W, Howell SH (1992) A single genetic locus, CKRl, defines Arabidopsis mutants in which root growth is resistant to low concentrations of cytokinin. Plant Physiol 99: 1569-1574
Teyssendier de la Serve B, Axelos M, Peaud-Lenoel C (1985) Cytokinins modulate the expression of genes encoding the protein of the light-harvesting chlorophyll a/b complex. Plant Mo1 Biol 5
Trewavas A (1983) 1s plant development regulated by changes in the concentration of growth substances or by changes in the sensitivity to growth substances? Trends Biochem Sci 8 354-357
Valvekens D, Van Montagu M, Van Lijsebettens M (1988) Agro- bacterium tumefuciens-mediated transformation of Arabidopsis thal- iann root explants by using kanamycin selection. Proc Natl Acad
Wang TL, Beutelmann P, Cove D (1981a) Cytokinin biosynthesis in mutants of the moss Physcomitrella patens. Plant Physiol 6 8 739-744
Wang TL, Horgan R, Cove D (1981b) Cytokinins from the moss Physcomitrella patens. Plant Physiol 6 8 735-738
pp 305-347
357-364
91: 5-8
155-163
Sci USA 8 5 5536-5540
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 1994 American Society of Plant Biologists. All rights reserved.





























