Embed Size (px)
Citation preview

A matrilin-2 expressziójának vizsgálata
májregenerációban kísérletesen és hepatocelluláris
carcinomában
Doktori értekezés
Szabó Erzsébet
Semmelweis Egyetem
Patológiai Tudományok Doktori Iskola
Témavezető: Prof Dr. Schaff Zsuzsa, egyetemi tanár, az orvostudományok doktora
Hivatalos bírálók: Dr. Mózes Miklós, egyetemi docens, PhD
Dr. Tóvári József, tudományos munkatárs, PhD
Szigorlati bizottság elnöke: Dr. Jeney András, egyetemi tanár,
az orvostudományok doktora
Szigorlati bizottság tagjai: Dr. Kovalszky Ilona, egyetemi tanár,
az orvostudományok doktora
Dr. Simon Károly, osztályvezető főorvos, PhD
Budapest 2007

1
TARTALOMJEGYZÉK
1. RÖVIDÍTÉSEK JEGYZÉKE........................................................................... 3
2. BEVEZETÉS ÉS IRODALMI HÁTTÉR .......................................................... 6
2.1. Hepatocelluláris carcinoma ................................................................................................................6 2.1.1. Epidemiológia................................................................................................................................6 2.1.2. Etiológia.........................................................................................................................................6 2.1.3. Hepatocarcinogenesis, a HBV és HCV fertőzés szerepe a patogenezisben...................................7 2.1.4. HCC típusai, lokalizációja, morfológiája......................................................................................8 2.1.5. A HCC szövettana .........................................................................................................................9 2.1.6. Májfibrosis, cirrhosis ...................................................................................................................10
2.2. Májregeneráció..................................................................................................................................11 2.2.1. A májregenerációban szerepet játszó sejtek, májregenerációs kísérleti modellek .......................11 2.2.2. A humán máj progenitor sejtjei ...................................................................................................14 2.2.3. Az őssejtek és a hepatocarcinogenesis kapcsolata .......................................................................15
2.3. Az extracelluláris mátrix (ECM).....................................................................................................18 2.3.1. Az ECM szerkezete és jelentősége ..............................................................................................18 2.3.2. Az ECM szerepe a tumorprogresszióban.....................................................................................19 2.3.3. ECM a normál májban.................................................................................................................20 2.3.4. A máj sejtjei, szerepük az ECM proteinek szintézisében.............................................................21 2.3.5. Az ECM változásai fibrosisban és cirrhosisban...........................................................................21 2.3.6. Az ECM jellemzői a májregeneráció során .................................................................................24
2.4. A matrilin fehérje család...................................................................................................................26 2.4.1. A matrilinek szerkezete ...............................................................................................................26 2.4.2. A család tagjainak jellemzése és filogenetikai eredete ................................................................28 2.4.3. A matrilinek oligomerizációja és kölcsönhatása más ECM komponensekkel.............................30 2.4.4. A matrilinek expressziója különböző szövetekben és szervekben...............................................33 2.4.5. A matrilinek szerepe betegségekben............................................................................................34 2.4.6. A matrilin-2 expressziója humán szövetekben és tumorokban....................................................36
3. CÉLKÍTŰZÉSEK.......................................................................................... 38
4. ANYAG ÉS MÓDSZER................................................................................ 39
4.1. A vizsgálatokhoz használt oldatok, pufferek összetétele ................................................................39
4.2. Állatkísérletek....................................................................................................................................40 4.2.1. Parciális hepatectomia (PH).........................................................................................................40 4.2.2. Parciális hepatectomia/acetylaminofluorén (PH/AAF) modell....................................................40
4.3. Humán szövetek.................................................................................................................................40
4.4. Morfológiai vizsgálatok.....................................................................................................................41 4.4.1. Hisztológia...................................................................................................................................41 4.4.2. Immunhisztokémiai vizsgálatok ..................................................................................................41
4.4.2.1. Paraffinos metszeteken végzett immunfestés.......................................................................41 4.4.2.2 Fagyasztott metszeteken végzett immunfloureszcenciás vizsgálat .......................................42 4.4.2.3. Fotódokumentáció................................................................................................................42 4.4.2.4. A matrilin-2 immunhisztokémiai reakciók morfometriai elemzése .....................................45

2
4.5. Molekuláris biológiai vizsgálatok.....................................................................................................45 4.5.1. Szöveti lézer mikrodisszekció és RNS izolálás patkány májból..................................................45 4.5.2. RNS-izolálás RNA laterben fixált humán anyagból ....................................................................45 4.5.3. cDNS előállítás ............................................................................................................................46 4.5.4. Primerek és a PCR körülményei ..................................................................................................47 4.5.5. Grádiens PCR ..............................................................................................................................47 4.5.6. Agaróz gélelektroforézis..............................................................................................................48 4.5.7. Real-time RT-PCR.......................................................................................................................49 4.5.8. Real-time RT-PCR eredmények elemzésének módszere.............................................................50 4.5.9. HCV vírus detektálása humán májmintákból Nested - PCR módszerrel .....................................51 4.5.10. RNS expresszió detektálása in situ hibridizációval ...................................................................55 4.5.11. Fehérje izolálás ..........................................................................................................................56
4.5.11.1. Patkány májakból...............................................................................................................56 4.5.11.2. Humán májakból................................................................................................................56
4.5.12. SDS-poliakrilamid gélelektroforézis .........................................................................................57 4.5.13. Western blot...............................................................................................................................57
5. EREDMÉNYEK ............................................................................................ 59
5.1. Matrilin-2 fehérje expresszió vizsgálata patkány májban immunhisztokémiával.......................59 5.1.1. Matrilin-2 vizsgálata immunhisztokémiával normál patkány májban .........................................59 5.1.2. Matrilin-2 immunhisztokémiai kimutatása májregenerációban..................................................60
5.2. Matrilin-2 fehérje expresszió detektálása AAF/PH kezelést követően Western blot technikával....................................................................................................................................................................66
5. 3. Matrilin-2-t termelő sejtek detektálása májregenerációban.........................................................68
5.4. Matrilin-2 expresszió vizsgálata real-time RT-PCR analízissel ....................................................70
5. 5. Matrilin-2 expresszió vizsgálata humán májban ...........................................................................71 5.5.1. Matrilin-2 fehérje expresszió vizsgálata ép és cirrhotikus humán májszövetben immunhisztokémiai módszerrel.............................................................................................................71 5.5.2. Matrilin-2 fehérje expressziójának vizsgálata hepatocelluláris carcinomában ............................72 5.5.3. A laminin és matrilin-2 kolokalizációjának vizsgálata humán májszövetben..............................74 5.5.4. Matrilin-2 immunhisztokémiai reakciók morfometriai elemzése és statisztikai analízise ...........75 5.5.5. Matrilin-2 fehérjeexpresszió vizsgálata Western blot technikával humán májakon ....................77 5.5.6. Matrilin-2 mRNS expressziójának real-time RT-PCR analízise humán májmintákon................78
6. MEGBESZÉLÉS .......................................................................................... 81
7. KÖVETKEZTETÉSEK ................................................................................. 88
8. ÖSSZEFOGLALÁS ..................................................................................... 89
9. SUMMARY................................................................................................... 90
10. IRODALOMJEGYZÉK ............................................................................... 91
11. SAJÁT KÖZLEMÉNYEK JEGYZÉKE ..................................................... 108
12. KÖSZÖNETNYÍLVÁNÍTÁS...................................................................... 112

3
1. RÖVIDÍTÉSEK JEGYZÉKE
AAF acetylaminofluorene, acetylaminofluorén
ABC avidin-biotin complex, avidin - biotin komplex
AEC 3-amino-9 ethyl-carbazole
AFB1 aflatoxin B1
AFP alpha-fetoprotein, alfa-fetoprotein
AS aminosav
BHMED bilateral hereditary microepiphyseal dysplasia, bilaterális örökletes
microepiphysealis dysplasia
BM basement membrane, bazális membrán
BMHSPG bazális membrán heparán szulfát proteoglikán
BSA bovine serum albumin, borjú szérum albumin
BT biotinilált tiramin
CC cholangiocarcinoma
cc coiled coil domén
CMP cartilage matrix protein, porc mátrix fehérje
COMP cartilage oligomeric matrix protein, porc oligomer mátrix fehérje
DAB 3,3’-diaminobenzidin tetrahidroklorid
DAPI 4',6-diamidino-2-phenylindol
ECL enhanced chemiluminescence, erősített kemilumineszcencia
ECM extracellular matrix, extracelluláris mátrix
EDTA ethylene -diamine-tetraacetic acid
EGF epidermal growth factor, epidermális növekedési faktor
ELISA enzyme linked immunosorbent assay, enzim-kötött immunoszorbens
próba
GAG glükózaminoglikán
GAPDH glyceraldehyde-3 phosphate dehydrogenase, glicerinaldehid-3-foszfát
dehidrogenáz
GBM multiform glioblastoma
HBV hepatitis B virus, hepatitisz B vírus
HCC hepatocellular carcinoma, hepatocellularis carcinoma
HCV hepatitis C virus, hepatitisz C vírus

4
HE hematoxilin –eozin
HRP horseradish peroxidase, tormaperoxidáz
HSPG heparánszulfát proteoglikán
IS idiopathic sclerosis, idiopatikus szklerózis
IL-1 interleukin-1
MDR-1 multidrog resistance gene-1, multidrog rezisztencia gén-1
MED multiple epiphiseal dysplasia, multiplex epipyseális dysplasia
MIDAS- metal ion dependent adhesion site, fémionfüggő adhéziós hely
MMP matrix metallpoproteinase, mátrix metalloproteináz
NF-κB nuclear factor-κB
NOH – AAF N-hidroxi- acetaminofluorén
ORF open reading frame, nyitott olvasási keret
PA pilocitikus astrocytoma
PBS phosphate buffer saline, foszfát puffer
PCR polymerase chain reaction, polimeráz láncreakció
PG proteoglikán
PH partial hepatectomy, parciális hepatectomia
PPARα peroxisome proliferator-activated receptor-α
PSACH pseudochondrodysplasia
RT reverse transcriptase, reverz transzkriptáz
SDS sodium dodecyl sulfate, nátrium dodecil-szulfát
SDS-PAGE sodium dodecyl sulfate-poliacrilamide gel electrophoresis, nátrium
dodecil-szulfát -poliakrilamid gél elektroforézis
SSC sodium chloride/ sodium citrate, nátrium-klorid/citrát
NTB nitro- tetrazolium-blue
BCIP 5-bromo-4-chloro-3-indolyl phosphate
TBE tris –borát- EDTA
TBS tris-buffer-saline, tris-puffer
TGF transforming growth factor, transzformáló növekedési faktor
TGF α transforming growth factor-α, transzformáló növekedési faktor- α
TGF β transforming growth factor-β, transzformáló növekedési faktor- β
TIF tubulointersticial fibrosis, tubulointerszticiális fibrosis (vesében)

5
TIMP tissue inhibitor metalloproteinase, szöveti metalloproteináz inhibitor
TNFα tumor necrosis factor-α, tumor nekrózis faktor-α
U unique domain, egyedi szekvencia
vWFA von Willebrand factor A, von Willebrand Faktor A

6
2. BEVEZETÉS ÉS IRODALMI HÁTTÉR
2.1. Hepatocelluláris carcinoma
A hepatocelluláris carcinoma (HCC) világviszonylatban az ötödik leggyakoribb
daganatos betegség. 2005-ben 667 000 esetet számoltak világszerte, ebből 17 500-at az
USA-ban (Rougier és mtsai 2007). Világviszonylatban évente körülbelül egymillió új
beteggel kell számolni, mortalitás tekintetében a harmadik helyen áll a malignus
daganatok között.
A daganat kezelése, különösen előrehaladott stádiumban megoldatlan, bár az utóbbi
hónapokban ismertetett cél-orientált gyógyszer, a sorafenib, talán áttörést jelent (Llovet
és mtsai 2007). A korai diagnózisban szerepet játszó érzékeny diagnosztikai markerek
feltárása, a posztoperatív szakban a HCC kiújulásának monitorozása változatlanul nagy
kihívást jelent.
2.1.1. Epidemiológia
A hepatocelluláris carcinoma (HCC) a humán carcinomák 5,6%-áért (férfiak: 7,5%,
nők: 3,5%) felelős. Prevalenciája az utóbbi években Európában és az USA-ban is
jelentősen növekedett, így a következő két évtizedben különösen jelentős orvosi és
népegészségügyi probléma lesz, amint az Japánban már ma is látható (Rougier és mtsai
2007). Magyarországon nem tartozik a leggyakoribb daganatok közé, de előfordulása
növekvő tendenciát mutat. (Szabo és mtsai 2003, Nagy és mtsai 2006).
2.1.2. Etiológia
A HCC etiológiájában több tényező játszik szerepet. Ezek között kiemelten a
vírusfertőzések, elsősorban a hepatitisz B vírus (HBV) és a hepatitisz C vírus (HCV)
fertőzés áll. A HCC előfordulásában kifejezett geográfiai különbségek tapasztalhatók.
Jellegzetes földrajzi eloszlást mutat, igen gyakori Afrika középső és déli területén,
Délkelet-Ázsiában, míg Európában és Észak-Amerikában ritkábban fordul elő (Buendia
1992, 2000). Az afrikai és az ázsiai HCC esetek többsége HBV fertőzéssel, míg a

7
Japánban, Európában és az USA-ban előforduló esetek HCV fertőzéssel függnek össze
(Hayashi és mtsai 1999, Aizawa és mtsai 2000, Szabo és mtsai 2003, 2004). A
legjelentősebb kofaktorok között az alkohol, az aflatoxin (AFB1), a dohányzás, egyes
szteroidok és egyéb toxikus tényezők szerepelhetnek a HCC etiológiájában. További
kockázati tényezőt jelentenek egyes genetikai rendellenességek (heamochromatosis,
Wilson kór, az α1-antitrypsin hiány), amelyek növelik a HCC kialakulásának esélyeit
(Hofseth és mtsai 2003). A cirrhosis önmagában is kockázati tényezőként jön
számításba, ugyanis a HCC-k 70-80%-a cirrhotikus talajon alakul ki. Újabban az
irodalom a nem alkoholos zsírmájat is kockázati tényezőként említi (Teufel és mtsai
2007).
Hazánkban elsősorban a hepatitisz C, majd a B és D, valamint az alkoholos cirrhosis és
a haemochromatosis a legfőbb oki tényező. A leggyakoribb HCV genotípus hazánkban
az 1b altípus (Gervain és mtsai 2001, Lengyel és Tulassay 2007).
2.1.3. Hepatocarcinogenesis, a HBV és HCV fertőzés szerepe a patogenezisben.
A HCC kialakulása többtényezős, többlépcsős és összetett folyamat, amely gyakran a
hepatitisz vírusok (HBV és HCV) perzisztens jelenlétével van összefüggésben. A HCC-
t tumorképződés számos jellemzője közül a gyors növekedés és infiltráció, a korai
metasztázisképzés, a nagyfokú malignitás jellemzi. A HCV-vagy a HBV-fertőzés lassan
progrediáló krónikus májbetegséget okoz, mely megjelenhet enyhe vagy súlyosabb
aktivitású krónikus hepatitis, cirrhosis vagy HCC formájában. A kiterjedt kutatások
ellenére sem a HBV és még kevésbé a HCV okozta májrák keletkezési mechanizmusa
nem egyértelműen tisztázott (Schaff és mtsai 2004). Számos adat a gyulladás és a
hepatocita-regeneráció jelentőségét hangsúlyozza a patogenezisben (Chisari 2000,
Szabo és mtsai 2004). Más szerzők a virális antigének direkt carcinogén hatását emelik
ki (Andriasini és Barnabas 1999).
A HCC–ben szenvedő betegek 40%-a HBV-vel fertőzött, bár ez kiemelten Afrikában és
Távol-Keleten jellemző (Hofseth és mtsai 2002, Szabo és mtsai 2004, Teufel és mtsai
2007). A HBV sajátságai közül kiemelendő, hogy a vírus-DNS integrációja a sejt DNS-
ébe nem szükséges a vírus replikációja szempontjából, azonban e mechanizmus
lehetővé teszi a virális genom perzisztálását a sejtben. Az integrálódott virális DNS a
hepatocita-proliferáció során a replikálódó sejt-DNS-sel együtt szaporodik. A HBV-

8
DNS-integráció többféle hatású lehet a sejtre; így kromoszomális DNS-instabilitást,
transzlokációt, duplikációt, kromoszóma rekombinációt, mutagenesist okozhat,
valamint inkomplett virális fehérjéket kódolhat. Ezek celluláris szabályozó gének,
tumorszupresszor gének, többek közt a p53 elvesztését eredményezhetik (Schaff és
mtsai 1995) Gyakori a p53 és más szupresszor gén mutációja, egyes növekedési
faktorok expressziója pedig fokozódik (Kiss és mtsai 2002). A vírus géntermékeinek,
főleg az X gén kódolta fehérjének a – HBX-nek szerepe van a HBV daganat-kialakulást
elősegítő hatásában. Az X gén p53 és RB fehérjékre kifejezett közvetlen hatását is
feltételezik (Staib és mtsai 2003).
A HCV fertőzéssel asszociált hepatocarcinogenesis folyamata kevésbé ismert mint a
HBV-vel asszociálté. A Flaviviridae családjába tartozó HCV egyszálú RNS vírus
(Clarke és mtsai 1997) nem integrálódik a fertőzött sejt genomjába, mivel nem
rendelkezik reverz transzkriptáz (RT) aktivitással. A vírusgenom strukturális (core,
burok) és nem strukturális fehérjéket kódol, amelyek enzimfunkciókkal bírnak- helikáz,
proteináz (NS3) vagy RNS-polimeráz (NS5) aktivitásuk van. A HCV fertőzés és a
dagnatképződés szempontjából a HCV sejtproliferációt moduláló hatása kap elsősorban
szerepet. Ezért a hatásáért egyes vírusproteinek, elsősorban a core-komponens, tehetők
felelőssé. Ugyanis a HCV core-fehérje, mint különböző virális és celluláris gén
regulátora befolyásolja egyes gének expresszióját, fokozott növekedési stimulust
okozva, mely elősegíti a HCC kialakulását (Barba és mtsai 1997). A core fehérje
közvetett módon aktiválja a TNF-α receptorát, a Raf-1 kináz, és az NF-κB útvonalakat,
amely a TNF-α -indukált és Fas-mediált apoptózishoz vezet (You és mtsai 1999, Block
és mtsai 2003). Az utóbbi években egyre nagyobb szerepet tulajdonítanak a NF-κB-
nukláris faktornak a máj carcinogenesisében (Arsura és Cavin 2005).
2.1.4. HCC típusai, lokalizációja, morfológiája
A HCC kialakulhat cirrhosis talaján és cirrhosismentes májban, az előző lényegesen
gyakoribb. A HCC megjelenési formái igen változatosak lehetnek, így makroszkóposan
szoliter, multiplex noduláris vagy diffúz formában mutatkozhat (Schaff és Nagy 1997).
Magyarországon a HCC túlnyomórészt (70-80%-ban) cirrhosis talaján alakul ki. A
kisebb méretű HCC-k tokkal rendelkezhetnek. A progresszió mértékét gyakran a
makroszkóposan is szembetűnő ún. fiókdaganatok megjelenése, valamint a vénák

9
inváziója jellemzi. A tumor színe többnyire szürkésfehér vagy bevérzés miatt vöröses,
néha az epetermelés miatt enyhén barnás-zöldes színű.
2.1.5. A HCC szövettana
Szövettanilag a HCC különböző szerkezetet és fokozatot mutathat (Schaff és mtsai
2004) Leggyakrabban trabeculáris, tubuláris vagy alveoláris formában jelentkezik.
Sajátos formák is ismeretesek, pl. a világossejtes („hipernefroid”), a pleomorf és a
fibrosus alak. Külön entitás a fibrolamelláris típusú HCC. Ezen jellegzetes szöveti
szerkezetű daganat cirrhosis mentes májban, fiatalabb életkorban fordul elő (átlagos
életkor 23 év).
A HCC TNM klasszifikációjának elve hasonló egyéb tumorokéhoz. A T1-T3 kategóriák
közötti differenciálást a 2 cm-es tumorátmérő, a daganat szoliter vagy multiplex volta, a
lebenyek érintettsége határozza meg. A HCC differenciáltsági fokozatai követik az I-IV
differenciáltsági fok szerinti beosztást (grade), a magasan differenciált formától a
differenciálatlan formáig (Schaff és mtsai 2004).
A HCC prognózisa rossz, ezért létfontosságú a korai diagnózis. Klinikai detektálásában
a szérum alfa-fetoprotein (AFP) -meghatározásnak nagy jelentőséget tulajdonítanak.
Noha az AFP hatékony markernek bizonyult a HCC monitorozásában és hisztológiailag
is detektálható, csak bizonyos korlátokkal alkalmazható. A korai HCC diagnózisában az
AFP mérése akár 40%-os hibát is okozhat (Deng-Fu Yao és mtsai 2007). Az AFP-
emelkedése a szérumban nem HCC – specifikus, mert más daganatokban (csírasejtes
tumorok, pancreatoblastoma stb.) ugyancsak emelkedett értéket mutathat. A recidívák
észlelésére azonban jól használható, valamint a cirrhosisban szenvedő betegek
követésére is lehetőséget nyújt. Újabb adatok szerint az AFP-termelés alapján a HCC-k
két jól elhatárolható csoportba sorolhatók (Lee és Thorgeirsson 2006). Ez a daganatok
differenciáltságával is összefügg: a magasabban differenciált daganatokban az AFP
mRNS expressziója alacsonyabb, mint a rosszul differenciáltakban (Craig 1997).
A HCC extrahepaticus terjedése elsősorban a perihepatikus és abdominális
nyirokcsomókba, a tüdőbe és a csontokba történik (Katyal és mtsai 2000, Schaff és
mtsai 2004).

10
2.1.6. Májfibrosis, cirrhosis
A máj krónikus elváltozásainak közös jellemzője, hogy többnyire fokozott kötőszövet
termeléssel járnak és májfibrosishoz vezetnek. A májfibrosis során az extracelluláris
mátrix kórosan felhalmozódik a májban, és szerkezeti átépüléshez vezethet (Schaff és
Illyés 2004, Kovalszky és mtsai 2006). Cirrhosis esetén a máj állománya nodulusosan
átépül, az eredeti acináris struktúra nem ismerhető fel. Diffúz, a máj szerkezetének
torzulásával, átépülésével, göbök kialakulásával és progresszív fibrosissal járó folyamat.
A májcirrhosis során a májállományt képező regeneratív göböket (nodulusok)
kötőszöveti rostokból álló sövények (septumok) vesznek köröl. A rostok között
limfocitás infiltráció és epeút-proliferáció észlelhető. A cirrhosis osztályozása a
morfológiai megjelenés és kórokok alapján történhet. Etiológiai szempontból a cirrhosis
legfontosabb formái: alkoholos, virális, biliáris, toxikus eredetű, posztnekrotikus,
autoimmun, metabolikus zavar okozta, egyéb fertőző ágens okozta és kriptogén (Schaff
és Illyés 2004).
A cirrhosis patogenezisében több tényező játszik szerepet. Ez egy multifaktoriális
folyamat, elindítója a sejtkárosodás, mely részben különböző immunológiai
mechanizmusok beindulását, másrészről a széteső sejtekből származó faktorok
felszabadulását idézi elő. A legfontosabb patogenetikai tényezők között a sejtpusztulás
eredményezte krónikus gyulladás, s ennek következtében különböző gyulladásos
citokinek (TNFα, TGFα, TGFβ, interleukinek) fokozott termelődése játszik szerepet. Ez
a különböző mezenhimális sejtek (Kupffer-sejt, endothelsejt, fibroblast stb.) aktiválását
és az extracelluláris mátrix különböző komponenseinek felhalmozódását eredményezi
(erről bővebben az Extracelluláris mátrix fejezetben lesz szó).

11
2.2. Májregeneráció
2.2.1. A májregenerációban szerepet játszó sejtek, májregenerációs kísérleti
modellek
Az egészséges felnőtt májban a sejtpusztulás és sejtképződés egyensúlyban van
egymással. A máj óriási regenerálódó képességgel bír (Sarraf és mtsai 1994, Taub
1996). Az elmúlt évek kutatásai során a legfontosabb kérdések egyike az volt, milyen
eredetű sejtekből történik a máj regenerációja a máj sérülését követően.
A máj regenerációban szerepet játszó folyamatok tanulmányozása állatmodelleken vált
lehetővé. Így az eddig felhalmozódott ismeretek elsősorban patkány májra vonatkoznak.
Normál körülmények közt a májsejtek minimálisan proliferálnak. A máj sérülését
követően a hepatociták a sejtciklus Go fázisába lépve regenerálják a sérült májszövetet.
Így a különböző májsejt - vesztéssel előidézett májsérülés, mint a parciális
hepatectomia, vírus fertőzés vagy más faktor okozta gyulladásos folyamatot követően a
májszövet újjáépülése elsősorban a hepatociták replikációjával történik (Forbes és mtsai
2002). Különböző állatmodellekbe transzplantált hepatocitákkal végzett kísérletek során
igazolódott, hogy a beültetett hepatociták bizonyos hányada klonális expanzióban vesz
részt, és jelentős regenerálódó kapacitással bír (Forbes és mtsai 2002) Ennek alapján
feltételezhető, hogy a hepatociták önmagukban is a máj funkcionális őssejtjei. A
májregeneráció hepatociták részvételével történő tanulmányozására általában a Higgins
és Anderson által már 1931-ben leírt parciális hepatectomiát (PH) alkalmazzák.
Eszerint, ha a máj 70%-át parciális hepatectomiával eltávolítjuk, a májtömeg 7-10 nap
múlva regenerálja önmagát. Ennek molekuláris mechanizmusát számos kutatócsoport
tanulmányozta (Fausto 2001). Ha azonban a májat súlyos károsodás éri és a hepatociták
sérülnek vagy replikációjuk gátolt, az ún. potenciális őssejtek (potencial stem cell)
kompartmentek aktiválódnak (Roskams és mtsai 2003, Fausto 2004, Santoni-Rugiu és
mtsai 2005). Ezek a bipotenciális képességgel bíró, más néven ovális sejtek, mind
hepatocitákká, mind epeút hámsejtekké képesek differenciálódni. Az ovális sejtek
jellemző epeúti markerei a CK7, a CK19, OV-6, OV-1, hepatocita markere az AFP,
neuroendokrin markerei a chromogranin-A, connexin 43, CD34 stb. (Roskams 2006).
Az ovális sejt elnevezést az ovális mag és a halvány bazofil citoplazma miatt kapta
(Farber 1956). A patkány májban a Hering canalisokat felépítő sejtek viselkednek

12
őssejtekként, melyekből aktivizálódásukkor tubulusszerű képleteket formáló ovális
sejtek képződnek (Paku és mtsai 2001, 2004). Ezek a tubulusszerű képletek
szövettanilag a Hering canalisok folytatását képezik.
Feltételezhetően az őssejteknek van egy harmadik populációja is, amely májsejt
potenciállal rendelkezik, és a csontvelőből származik. Ezek a heamatopoietikus őssejtek.
Jóllehet ezeknek a sejteknek a megújulási sebessége viszonylag alacsony,
jelentőségteljes szerepük felmerül a regenerációban (Theise és mtsai 2000, Alison
2005).
Noha az utóbbi években igen kibővült az őssejtek eredetével fogalkozó irodalom,
egészen pontosan még a mai napig sem derült fény ezek pontos anatómiai
lokalizációjára, növekedési mintázatára. Az azonban ismert, hogy ezek az őssejtek a
periportális területen proliferálnak és innen terjeszkednek a májparencymába. Ezt az is
bizonyítja, hogy a periportális régió szelektív roncsolásával jelentősen csökkent a
proliferáló ovális sejtek száma (Petersen és mtsai 1998). A Hering canalisból való
eredetüket az ovális sejtek és az epeutak hámsejtjei közötti igen kifejezett fenotípusos
hasonlatosság is alátámasztja. A Hering canalisok kötik össze a disztális hepatocitákat
az interlobuláris epeúti duktuszokkal, amelyek valószínűleg az őssejtek
„szálláshelyei”(Sell és Salman 1984). Egyes szerzők szerint valamennyi epeút hámsejt
potenciálisan képes őssejtként viselkedni, míg a hepatocitákra jellemző transzkripciós
faktorok megjelenése az ovális sejtekben hepatocita irányú differenciálódást jelezhet.
Az 1. ábra a tudomány jelenlegi álláspontja szerinti ovális sejtekre vonatkozó
terminológiát és lokalizációjukat tünteti fel.

13
1. ábra. Az őssejtek lokalizációja a májban (Roskams 2006).
A bal oldali rész a portális teret (PT) ábrázolja. Az epeút (biliary tree) legkisebb és
legtávolabbi elágazódása portális biliáris duktuszokból (BD) és intralobuláris
duktuszokból áll. A Hering canalisokat részben hepatociták, részben
cholangiocyták képezik. Az őssejtek a Hering canalis legtávolabbi elágazódásai. A-
artéria, V- centrális véna.
Wilson és Leduc (1958) írták le először a tartalék „őssejt kompartmentet” súlyos diétát
követően egérben, melyet azóta patkányban több kísérleti modellben is sikerült
kimutatni. A legtöbb modellben parciális hepatectomiát követően egy carcinogén
vegyülettel gátolják a májsejt proliferációt.
Az ovális sejt kompartmentek indukálta májregeneráció vizsgálatára a 2-
acetylaminofluorén (AAF)/parciális hepatectomia (PH) modell az egyik legalkalmasabb
kísérleti rendszer (Evarts és mtsai 1987, Nagy 1995).
Kísérletileg igazolták, hogy már az AAF kezelést követően, a hepatectomia előtti
periódusban is a májban jól definiált változások figyelhetők meg. Egyetlen AAF dózis
adagolása után is szignifikánsan megemelkedik a periportális régióban az osztódó sejtek
száma (Paku és mtsai 2001, 2004). Az acetylaminofluorén a fluorénből származó
carcinogén vegyület. A fluorént a szervezet nem metabolizálja, ezért az egy ártalmatlan,
carcinogén hatással nem rendelkező anyag. Az aminofluorént (AF) már képesek a
májsejtek acetaminofluorénné átalakítani, amiből N- hidroxilációval alakul ki a
carcinogén hatású, DNS-hez kötődni képes N-hidroxi-acetilaminofluorén (NOH AAF).

14
Az AAF a májsejtek DNS molekuláihoz való kötődése révén megakadályozza azok
belépését a sejtciklusba. Az AAF kezelést követő PH után az ovális sejtek proliferálnak
és messze a májparenchymába terjednek a májsejtgerendák közé, ott hepatocitákká
differenciálódnak. Az AAF kezelés parciális hepatectomiával történő kombinációja
során a proliferáló ovális sejtek sokkal nagyobb számban figyelhetők meg. Az ovális
sejtek hepatocitává történő differenciálódásában döntő fontosságú esemény, amikor az
ovális sejtekből „bazofil fókuszok” formálódnak. A fókuszokat felépítő sejtek
morfológiailag is átmenetet mutatnak az ovális sejtek és a hepatociták között, mind
nagyság, mind alak vonatkozásában. Funkcionálisan is a fókuszokat felépítő sejtekben
jelennek meg a magasan differenciált hepatocitákra jellemző vonások (jelentősen
fokozott albumintermelés, csökkent AFP expresszió, MDR-1 expressziója, HNF-4)
(Nagy 1995, Santoni-Rugiu és mtsai 2005).
Az AAF analóg vegyületek is képesek a periportális sejproliferáció kiváltására.
Dexamethason gátolja az ovális sejtek és a hepatociták osztódását is parciális
hepatectomiát követően. Ha a májregeneráció mindkét mechanizmusát - hepatocita
proliferáció, őssejt aktivizálás - gátolták, a máj tömege a hepatociták hypertrophiája
révén pótlódott. A periportáis zónában ilyenkor a hepatociták jelentős
megnagyobbodása figyelhető meg (Nagy és mtsai 2001). Az AAF-hez hasonló hatást
lehet elérni szén-tetraklorid (CCL4) adagolásával vagy ethionin intoxikációval is
(Alison és mtsai 1996). Az egérben előidézett zsírmáj modellekben, mint például az
ObOb (-/-) egérben vagy a PARP (-/-) egérben is gátolt a hepatociták proliferációja és
az ovális sejtek aktiválása jellemző (Roskams és mtsai 2003).
2.2.2. A humán máj progenitor sejtjei
Az eddigi irodalmi adatok alapján úgy tűnik, hogy a típusos ovális sejtek a patkány
májra jellemzőek. Egérben jóval nehezebben sikerült hasonló sejteket indukálni.
Hasonló a helyzet a humán májban is. Az emberi májszövetben az ovális sejt
proliferációhoz egy hasonló jelenséget, az un „atipusos duktuláris proliferációt” lehetett
megfigyelni elsősorban epeút elzáródással járó különböző májbetegségekben (Roskams
2006). Ezek az ovális sejtektől eltérően azonban nem mutatnak AFP expressziót.
Feltételezhető, hogy az atípusos duktuláris reakciók az emberi májban egy, az
őssejtekből kiinduló regeneratív válasznak felelhetnek meg. Számos morfológiai

15
hasonlóságot véltek felfedezni az ovális sejtek és a humán májban az ún. duktuláris
reakciók között:
• biliáris típusú citokeratinokat expresszálnak
• a periportális zónából a májlebenybe terjeszkednek
• közeli kontaktusban vannak az aktivált csillag sejtekkel
• növekedésüket ugyanaz a növekedési faktor rendszer szabályozza
• képesek hepatocitákká differenciálódni
Néhány közlemény óvakodik a duktuláris fenotípus elnevezéstől, így inkább IMHBC
(„intermediate hepatobilliary cells”) –névvel illeti a közepes méret (6 µm <IMHBC <40
µm), a CK19, CK7 pozitivitás és egyes hepatocita markerek (albumin, α1-antitrypsin,
HepPar1) pozitivitása miatt. Feltétlenül meg kell különböztessük a duktuláris reakciót
az abnormális non-reaktiv epeútszerű képződményektől („ductal plate malformations”),
pl. a fibropolycystikus májbetegségekben (Desmet 2005).
A máj különböző elváltozásaiban figyeltek meg duktuláris reakciót, amelyet az alábbi
csoportokba sorolhatunk:
A) A máj károsodásával lejátszódó duktuláris reakció (pl. masszív májnekrózis, súlyos
zsíros degeneráció, cirrhosis stb);
B) A máj károsodása nélkül lezajló duktuláris reakció (pl. krónikus hepatitisz, alkoholos
májbetegség, fokális noduláris hyperplasia (FNH), hosszan tartó hideg ischémia stb);
C) Májtumorokban (pl. peritumorális duktuláris reakció, köztes fenotípusú tumorokban)
(Thung 1990, Hsia és Evarts1992).
A duktuláris reakció intenzitása korrelál a gyulladásos folyamat intenzitásával és
hozzájárulhat a krónikus heptitisz során lejátszódó fibrogenezishez (Eleazar és mtsai
2004). Jó néhány olyan esetet közöltek már az irodalomban, amikor májnekrózist
követően a máj regenerálódása ezeknek a duktuláris sejteknek a részvételével zajlott
(Fujita és mtsai 2000).
2.2.3. Az őssejtek és a hepatocarcinogenesis kapcsolata
Még a 19. században Virchow figyelte meg, hogy számos tumorra jellemző az
embrionális eredet (Sell 2004). A tumorok az őssejtek malignus transzformációjából is
származhatnak. Az ovális sejtek hepatocarcinogenesisben betöltött szerepének direkt
bizonyítékát Dumble írta le (2002), amikor p53-mutáns egerekből izolált ovális sejteket

16
thymus-sérült egerekbe oltva HCC-t generált. Jelenleg a májban előforduló hosszú életű
heptocitákat, a cholangiocytákat és a bipotenciális őssejteket (amelyek a Hering
canalisok legtávolabbi részéből származnak) tekinti az irodalom a májban lezajló
carcinogenesis legújabb célpontjainak (Roskams 2004). Néhány krónikus
májbetegségben kimutatták, hogy a telomerhossz fokozatos rövidülése miatt (ezt igen
jellemzően figyelték meg cirrhotikus májban) a hepatociták „replikációs képessége
elöregszik”. A krónikus gyulladás, a növekedési faktorok jelenléte, a DNS-károsító
ágensek és a folyamatos replikáció egyaránt szerepet játszhat ebben a kórfolyamatban.
Ez a jelenség indukálja az őssejteket (a duktuláris reakciót) humán májban is. Krónikus
hepatitisz C-ben a duktuláris reakció intenzitása és a fibrosis mértéke között jelentős
összefüggés van (Lowes és mtsai 1999). A máj gyulladásos megbetegedéseiben a
hepatocitáknak egy olyan átmeneti alakját figyelték meg, amelyek átmenetet képeznek
az őssejtek és a hepatociták között. Ezek száma összefüggésben van a gyulladás
mértékével és a nekrózis fokával. Mindezek alapján valószínű, hogy ilyenkor elsősorban
az őssejtek differenciálódnak hepatocitákká, ezért nő meg ezen átmeneti hepatocita
alakok száma. Megfigyelték, hogy cirrhotikus májban és főképpen nem alkoholos
szteatohepatitiszben, a cirrhotikus nodulusokat is ilyen átmeneti hepatocita alakok
képezik. Igazolták, hogy ezek az ún. „hepatocita rügyek”, minden esetben reaktív
duktuszokkal vannak kapcsolatban. Ez is alátámasztja az őssejt eredetüket (Falkowski
2003).
Számos tanulmányban írták le a májtumorok monoklonalitását. A májra jellemző
legfontosabb rosszindulatú daganatoknak, a HCC-nek, a cholangiocelluláris
carcinomának (CC) és a kevert típusuknak (HCC/CC) eredete még máig sem teljesen
tisztázott. A HCC és CC fokális prekurzor léziókból alakul ki, egy többlépcsős folyamat
eredményeként, amelynek az őssejtek aktív résztvevői. Ezt az is bizonyítja, hogy a
HCC-k nagy része expresszálja az őssejtekre jellemző markerek egy részét (CK7,
CK19, OV6). A tumorsejtek ezekben a daganatokban fenotípusosan átmeneti hepatocita
alakokból és éretlen hepatocitákból állnak.
A tumorokban az őssejtek jelenlétét elsősorban két elmélettel hozzák összefüggésbe
(Roskams és mtsai 2006):
1. Az egyik szerint a tumorsejtek őssejt eredetűek („maturation arrest theory”)

17
2. A második szerint a tumor sejtek dedifferenciálódnak és progenitor sejt
tulajdonságokat vesznek fel a carcinogenesis során
Ez utóbbival magyarázható az, hogy a kis sejtes diszplasztikus fókuszok (< 1nm) 55%-a
progenitor sejtekből és átmeneti hepatocita alakokból áll. Míg a nagy sejtes
diszplasztikus fókuszokat érett pusztuló hepatociták alkotják.
CC-ben is sikerült azonosítani egyes őssejt markereket. A hepatoblastomában (HB), egy
igen gyakori gyerekkori tumorban is kimutatták az ovális sejteket. Ezt a daganatot őssejt
eredetűnek tartják (Fiegel és mtsai 2004).
Összességében tehát minden olyan krónikus májbetegségben, amely kockázati tényezőt
jelent a HCC vagy CC kialakulásához, az őssejtek (progenitor sejtek) proliferálnak, a
hepatociták pedig szeneszcensé válnak (Roskams 2004).
Az őssejt eredetet és a hepatocarcinogenesis összefüggését tumorokban a 2. ábra
szemlélteti.
2. ábra. Az őssejtből kiinduló lehetséges májdaganatok. Az őssejtek mind
hepatocitákká mind cholangiocytákká képesek differenciálódni (Roskams 2006).

18
2.3. Az extracelluláris mátrix (ECM)
2.3.1. Az ECM szerkezete és jelentősége
A szövetek felépítésében nemcsak sejtek, hanem az ezeken kívül található, az
extracelluláris teret kitöltő makromolekulákból (fehérjékből és szénhidrátokból) álló
hálózat – az ECM is részt vesz. A mátrixot felépítő molekulák sokfélék lehetnek, a
szövet által betöltött feladatnak megfelelően. Kémiai természetük alapján a következő
nagy kategóriákba sorolhatók a mátrix molekulái (Aumailley és Gayraud 1998,
Kovalszky 2004):
1. Nagy poliszacharid-láncokból felépülő glükózaminoglikánok (GAG) és
proteoglikánok (decorin, perlecan, versican, syndecan, agrin stb)
2. Fibrilláris strukturális fehérjék (kollagének, elasztin, fibrillin-1,-2, entactin stb)
3. Nem- kollagén szerkezetű fehérjék (laminin, fibronectin, vitronectin, tenascin,
entactin stb.).
3. ábra. Az ECM szerkezete. www. fasebj.org/cgi/content/full
proteoglikánok
Cukorláncok
kollagének
integrinek
fibronektin

19
Jelenleg a mátrixban előforduló kollagéneknek 26 típusát ismerjük (Gelse és mtsai
2003). Ezek főbb csoportjai a következők: fibrilláris kollagének (I-III, V, XI),
hálózatképző kollagének (IV, VIII, X), fibril- asszociált kollagének (IX, XII, XIV, XIX),
transzmembrán kollagén (XIII és XVII), stb.
A nem – kollagén szerkezetű ECM molekulák csoportjába szerkezetileg hosszú,
flexibilis, több kollagén típust kötő doménnel rendelkező molekulák sorolandók,
melyek más mátrix proteineket, poliszacharidokat, sejt-felszíni fehérjéket, növekedési
faktorokat és hormonokat képesek kötni. Kulcsfontosságú szerepük van a matix-matrix
és sejt-mátrix kapcsolatok kialakításában és fenntartásában. Ezek a multiadhézios
fehérjék a sejt-mátrix kapcsolatot az RGD domén részvételével valósítják meg. A
mátrixot felépítő molekuláknak ebbe a csoportjába tartozik a dolgozatom fő témája, a
nemrégiben leírt matrilin fehérje molekulák családja is, melyek képesek kötni a
kollagéneket és más ECM komponenseket (Deák és mtsai 1999) (erről majd a további
fejezetekben írok).
2.3.2. Az ECM szerepe a tumorprogresszióban
A daganatkutatás a mátrix (stroma) jelentőségét sokáig nem ismerte fel. A daganatokat
főleg genetikai szempontból vizsgálták és a fenotípus változással magyarázták a
tumorképződést. Mára már elfogadott tény, hogy a daganat kötőszövetes alapállománya
nagymértékben hozzájárul a daganatok kialakulásához. Ennek megfelelően az utóbbi
években az ECM a tumorbiológia egyik legintenzívebben kutatott területévé vált (Füle
és mtsai 2006, Li és mtsai 2007).
Az alapállományba hatolva a daganatsejtek úgy alakítják a mezenchimális sejtek
működését, hogy azok olyan mátrixot állítsanak elő, mely összességében elősegíti a
daganatsejtek további invázióját. A daganat növekedéséhez szükség van oxigénre és
tápanyagellátásra egyaránt, amelyet a daganat az érújdonképződés (neoangiogenesis)
segítségével old meg. A tumorok neoangiogenesisének szabályozásában szerepet játszó
számos tényező között (növekedési faktorok, sejtadhéziós molekulák és egyéb
mediátorok) meghatározó szerepet kapnak az ECM-molekulák is.

20
A tumorszövet kötőszöveti állományában jelen levő ECM mind mennyiségi mind
minőségi szempontból jelentősen eltér a normál szövetekben találtaktól.
Mennyiségi változások: A daganatos stromában több kollagén, proteoglikán és
glükózaminoglikán van jelen, mint az egészséges szövetben (Kalluri 2003). A heparán-
szulfát magasabb szintű felszaporodását például melanómában, májrákokban,
veserákokban figyelték meg. A laminin egyes alegységeinek nagymértékű
felszaporodását áttétképző melanómákban és fibrosarcomákban igazolták (Gianelli és
Antoniaci 2000, Canen és Yamada 2001, Chiquet- Ehrismann 2004).
Minőségi változások:
A daganatok ECM fehérjéi nemcsak mennyiségükben, hanem esetleg szerkezetükben is
eltérhetnek az egészséges sejtközötti állománytól. A nagy mennyiségben lerakódott
proteoglikánok általában módosultak. A fibronektin, tenascin, CD44 és versican
különböző formáit mutatták ki daganatokban (Kovalszky és mtsai 2004).
2.3.3. ECM a normál májban
Az ECM az egészséges májban részben a máj rostos vázát képezi, részben a Disse-
térben található. A normál májban relatív mennyisége alacsony, kevesebb mint 3% -ot
tesz ki. Strukturális szerepe mellett aktív szerepe van a májat alkotó sejtek
működésének szabályozásában és nélkülözhetetlen a hepatociták differenciált
funkciójának az ellátásához. A rostos mátrixot képező molekulák közt elsősorban a
kollagéneket (I, III, V, VI), a decorint és a fibronektint említhetjük meg. Eddigi
ismereteink szerint a bazális membrán (BM) felépítésében elsősorban a laminin, a IV
kollagén és a heparánszulfát PG-ok (BMHSPG) játszanak szerepet.
A rostos mátrix a periportális területen és centrális véna körül fordul elő, de elemei
finomabb módszerekkel elszórtan a Disse-térben is kimutathatók. Normál májban a BM
főleg periportálisan és a centrális vénák körül látható, de bizonyították, hogy az összes
BM-t alkotó fehérje megtalálható a Disse-térben is, bár nem szerveződnek klasszikus
értelemben vett BM-ná (Kovalszky 1993). A BM megjelenését a szinuszoidok mentén
fibrosisban és cirrhosisban detektálták (Kovalszky és mtsai 2006).

21
Összefoglalva, tehát a májban, klasszikus értelemben vett magas denzitású BM van az
erek körül, míg periszinuszoidálisan alacsony denzitású BM található.
2.3.4. A máj sejtjei, szerepük az ECM proteinek szintézisében
A hepatociták a magasan differenciált, máj specifikus működésében játszanak szerepet,
a mátrix fehérjék közül azonban feltehetően csak a szöveti fibronektin termelésére
képesek. A szinuszoidokat bélelő endothel sejtekben trombospondin, fibronectin,
kollagén IV, entactin, HSPG és decorin expresszióját mutatták ki, ami azt bizonyítja,
hogy az ECM fehérjéinek nagy része innen származik (Schaffener és Popper 1963,
Maher és McGuire 1990). A periszinuszoidális térben a hepatociták és endothel sejtek
között található Ito sejtek (“fat storing” sejtek vagy lipociták, vagy HSC-hepatic stellate
cells, csillag sejtek) figyelhetők meg, amelyek több irodalmi adat alapján a matrix
fehérjék legfontosabb termelői. A “pit” sejtek máj asszociált lymphocyták, a
szinuszoidok lumenébe vannak lehorgonyozva, szoros kapcsolatban az endothel és
Kupffer-sejtekkel. Pontos szerepük még nem ismert, főleg a fibrogenezist stimuláló
faktorok termelésében lehet szerepük (Friedman 2004a, b).
Az ECM-t termelő sejtek körül még mindig vita folyik. Még máig is az egyik leginkább
elfogadott elképzelés, mely szerint a Disse-térben található periszinuszoidális
csillagsejtek aktiválódnak, fenotípust váltanak, myofibroblasttá alakulnak át. Egy másik
feltevés, hogy legalább három különböző eredetű sejtpopuláció ölti fel a myofibroblast
fenotípust és termel kötőszövetet. Az irodalmi adatok elsősorban a periszinuszoidalis
csillagsejtekkel foglalkoznak, sokkal kevesebb adat áll rendelkezésre a portális
fibroblastokról. A legutóbbi álláspont (2005 decembere) szerint a periszinuszoidális
csillagsejtek, a portális és centrális véna körüli fibroblastok mellett még csontvelői
őssejt eredetű myofibroblastok is találhatók a májban (Kovalszky és mtsai 2006).
2.3.5. Az ECM változásai fibrosisban és cirrhosisban.
A fibrotikus máj ECM komponensei nagymértékben megegyeznek a normál máj ECM
komponenseivel, azonban mennyiségi szempontból jelentősen megváltozik az ECM a
fibrotikus és cirrhotikus májban. A fibrosisban a lefontosabb esemény a mátrix jelentős
átépülése során bekövetkező változás, de ez a folyamat szoros összefüggésben áll olyan

22
történésekkel, mint a például a májsejt regeneráció, az érátrendeződés („vascular
redistribution”). Mindezek igen jelentősen befolyásolják a máj funkcióját. Noha a
krónikus májbetegségekben a kötőszövet elsősorban a portális területeken szaporodik
fel, a máj funkciója szempontjából nem ez, hanem a periszinuszoidális régió
megváltozása a kritikus. A fibrosis, cirrhosis során a szinuszoidok kapillarizációja a
legfontosabb kísérőjelenség. Ilyenkor az endothel sejtek elvesztik fenesztráltságukat, a
szinuszoidok helyett kapillárisok alakulnak ki. Az endothel és a hepatociták között az
alacsony denzitású BM helyét, magas denzitású BM foglalja el. Ennek eredményeként a
szabad, kétirányú molekulaáramlás, amely a plazmatér és a hepatociták közt fennáll
egészséges májban, akadályozva van. A májsejtek méregtelenítési és intermedier
anyagcserében betöltött szerepüket képtelenek ellátni (Martinez-Hernández és mtsai
1991).
A kötőszövet mennyiségét a májban a termelés és lebontás egyensúlya szabja meg, így
aktuális mennyisége a képződés és lebontás eredője. A kötőszövetet a kollagenázok
bontják le, melyek termeléséért szintén a már említett myofibroblastok tehetők
felelőssé. A kollagenázokat gátló TIMP (metalloproteáz-gátló) és α2 macroglobulin
fokozott termelése is az ECM felszaporodását idézi elő. A 4. ábra a fibrosisban és
cirrhosisban az ECM átalakulásához és a mátrix molekulák felfalmozódásához vezető
fibrogenezis kaszkádot mutatja be.

23
hepatocita károsodás
TIMP α2 macroglobulin
nonparenchymális májsejtek aktiválása
endothel, Kupffer
Kötőszövet produkciót fokozó citokinek termelése
TGFβ, TGFα, IL1, EGF, bFGF, TNFα
Májkárosítás
hepatocita pusztulás
myofibroblastMononuclearis
sejtek aktiválódása
csillag sejt , portális fibroblast, csontvelői őssejt
aktiválása
ECM termelése (kollagén, laminin,
fibronektin, proteoglikán)
Kötőszövet + -
kollagenázok
4. ábra. A májfibrosis kialakulásának eseményei (Kovalszky és mtsai 2006 alapján).

24
2.3.6. Az ECM jellemzői a májregeneráció során
Egészen mostanáig nagyon keveset tudunk az ECM átalakulásáról a máj regenerálódás
során. Még ma sem tisztázott teljesen, hogy milyen mechanizmusok döntik el az akut
vagy krónikus máj sérülés kimenetelét: az előbbi májregenerálódást, az utóbbi cirrhosist
indukál.
Egy lényeges eltérés azonban biztos: a májregenerálódás során a szinuszoidok
újraképződése, míg cirrhosisban a kapillarizáció a jellemző, amely magas denzitású BM
képződéssel jár.
A máj kitűnően alkalmazható az ECM tanulmányozására, mivel nemcsakhogy
„sebgyógyulással” reagál, mint más szervek, hanem regenerálódik is. E folyamat során
nemcsak sejtproliferáció játszódik le, de az ECM-nek egy jelentős felhalmozása és
átalakulása is végbemegy. A legfontosabb ECM komponenseket: fibronektin, laminin
kollagén (I, III, IV) több tanulmányban is vizsgálták (Martinez-Hernandez és mtsai
1991, Martinez-Hernandez és Amenta 1993). Számos eltérést figyeltek meg a
májregenerációban, a cirrhosisban és a fibrosisban bekövetkező ECM átépülésében.
Kim és mtsai (1997) a fibronektin emelkedett, majd csökkent expresszióját írta le a
Disse-térben a májregeneráció első napjaiban, amit azzal magyaráztak, hogy a
fibronektin eltávolítása az ECM-ből a hepatociták osztódásának előfeltétele. Ezzel
szemben krónikus májsérülést követően fellépő cirrhosis során a fibronektin igen korai
depozíciója figyelhető meg a Disse-térben. Feltehetőleg ez a fehérje, mint az ECM
szerveződését segítő mátrix komponens funkcionál. Ugyancsak eltérést figyeltek meg a
laminin és entaktin szekréciójában is. A cirrhosisban megjelenő entaktin és laminin a
szinuszoidok kapillarizációja során képződő BM kialakításában tölthet be fontos
szerepet (Martinez – Hernandez és Amenta 1995).
A regenerálódó máj kollagén tartalmában is hasonló csökkenést észleltek a
cirrhotikushoz képest. Összességében véve a májregenerációban tehát az ECM/sejt
tömeg csökkenés figyelhető meg. Mire a regenerálódás során a hepatociták
vaszkularizációja megtörténik, és kialakulnak a szinuszoidok ez az arány
normalizálódik.
Közvetlenül PH-t követően a laminin, entactin és fibronektin szintje is csökken a
májban, majd később emelkedik a regeneráció során. Fokozott laminin expressziót

25
figyeltek meg a regenerálódó máj szinuszoidjaiban (Kim és mtsai 1997, Paku és mtsai
2001, 2004). A lamininnek a BM-ban betöltött szerepén túl igen jelentős szerepet
tulajdonítanak a növekedés stimulálásában, a sejt migráció modulálásában, a
sejtdifferenciációban és a morfogenezis eseményeiben, mely egy integrineket aktiváló
szignalizációs kaszkádon keresztül stimulálja a májsejt proliferációt.
Más nem- kollagén ECM mátrix komponensek szerepe, így például a tenascinok és
egyéb BM komponensek (kollagének) konkrét funkciója és változásai a
májregenerációban és a cirrhosisban nem egészen világos. Ez a kérdéskör a további
kutatások tárgyát képezi. Egyértelmű azonban, hogy a fentebb említett molekulákkal
kölcsönhatásban levő fehérjék szerepe is felmerül a májregenerációban és a máj
elváltozásaiban (ezek közt említendők a matrilinek is).

26
2.4. A matrilin fehérjecsalád
2.4.1. A matrilinek szerkezete
A család prototípusa a matrilin-1, mely korábban porc mátrix proteinként volt ismert
(CMP, cartilage matrix protein) (Kiss és mtsai, 1989). A porc mátrix fehérje a matrilin
család elsőként felfedezett tagja, kizárólag a porc érése során expreszálódik. Ezt a
fehérjét a porc különböző formáiban proteoglikánokkal asszociálva írták le. (Paulsson
és Heinegard 1979, 1981, 1982). A matrilin-2, matrilin-3 (Belluoccio és Trueb 1997,
Deák és mtsai 1997, Wagener és mtsai 1997, Belluoccio és mtsai 1998) és a matrilin-4
(Wagener és mtsai 1998a) felfedezésével derült fény egy közös szerkezeti szekvencia
felderítésére.
A matrilin fehérjecsalád a nem-kollagén ECM fehérjéknek abba az alcsaládjába
sorolandó, amelybe a von Willebrand A factor (vWFA) domént tartalmazó fehérjék
tartoznak (Whittaker és mtsai 2002). A vWFA domén számos extracelluláris és
intracelluláris fehérje molekulában szerepel és gyakran résztvevője azoknak a fehérje-
fehérje kölcsönhatásoknak, amelyek a multimer fehérjék kialakításában játszanak
szerepet. A vWFA domén 200 aminosav (AS) maradékból épül fel, amelyek a
klasszikus Rossman fold szerkezetet hordozzák, középen a β-lemezzel és körülötte az α-
helikális struktúrákkal. Minden matrilin vWFA doménjére jellemző a konzervatív
szerkezetű MIDAS (metal ion dependent adhesion site, fémionfüggő adhéziós hely)
motívum, amelynek kitüntetett ligandumkötő szerepe van. A matrilin fehérjék vWFA
doménje szerkezetileg egy egyedülálló csoportot alkot az ugyanezt a domént tartalmazó
ECM molekulák között: érdekes megfigyelés, hogy minden matrilin családtag sokkal
nagyobb szerkezeti hasonlóságot mutat egymással, mint minden más vWFA modult
tartalmazó molekulával. Ez egyértelműen egy közös filogenetikai ősre utal (Deák és
mtsai 1999). A filogenetikai vizsgálatok alapján a matrilinekre jellemző két vWFA
domént két különböző csoportba sorolták be, amely szerint a matrilin-1/-3 és a matrilin-
2/-4 két külön alcsoportot képez a családon belül (Deák és mtsai 1999). A matrilinekben
található vWFA domének az N-terminushoz közel eső vWFA1 domén és a C-
terminushoz közel eső vWFA2 domén. A matrilin-3-ban a vWFA2 domén és az egér
egyik splice-variáns matrilin-4-ben a vWFA1 domén kiesett.

27
A matrilin-1-et a csirke, egér és emberi cDNS-ből írták le. Szerkezetileg két vWFA
domént tartalmaz, amelyet egy darab EGF-szerű domén köt össze. A matrilin-1 és
matrilin-3 splice-variánsaiból, hiányzik az EGF-szerű domén. Az összes többi
családtagban legalább egy ilyen domén van, legtöbb belőle a matrilin-4 splice-
variánsában található, amely 12 EGF-t tartalmaz (Ko és mtsai 2005). Ez az EGF-szerű
domén nem mutat szerkezeti hasonlóságot a Ca2+- kötő EGF-szerű motívumokkal: nem
Ca2+- kötők és növekedési faktor aktivitásuk sem bizonyított. Nagyobb valószínűséggel
egy elosztóként (ún. „spacerként”) szerepelnek a vWFA domének között. Szerkezetileg
leginkább a fibulin, a fibrillin EGF doménjeivel mutatnak rokonságot.
Végül a molekulák karboxi végén egy úgynevezett coiled-coil (cc) struktúra
helyezkedik el, amely α-hélixet formál. Ez teszi alkalmassá az egyedülálló alegységeket
a virágcsokorszerű oligomerizációra (Argaves és mtsai 1987, Kiss és mtsai 1989,
Hauser és Paulsson 1994). Az összes többi matrilin családtag ugyanezeket a doméneket
tartalmazza és ugyanolyan sorrendben, mint a matrilin-1. Kivétel jellemzi a matrilin-3-t,
amelyik szerkezetét jellemzi, hogy csak egyetlen vWFA doménje van és az EGF-
domének közvetlenül a C-teminálison levő coiled-coil szakaszhoz kapcsolódnak.
Továbbá, a matrilin-2 és a matriln-3 a fent említett doméneken kívül az N-terminálison
pozitív töltésű oldalláncokkal bíró AS maradékokat hordoz. Egyedül a matrilin-2-t
jellemzi egy úgynevezett egyedi (U, unique) szekvencia, a második vWFA domén és az
oligomerizációs domén között, amely egyedi, eddig még egyetlen más fehérjében sem
írtak le hozzá hasonlót.
A humán matrilin-2 prekurzort 4.0 kb mRNS kódolja, 956 aminosavból áll és 93%-os
egyezést mutat az egér fehérjével. 2 vWFA domént, 10 EGF-domént, egy egyedi
szekvenciát és egy oligomerizációs domént tartalmaz. Az első vWFA domén a
legkonzerváltabb. A matrilin-2 a 8q22 kromoszómára lokalizálódik. Az 5. ábrán a
matrilin család domén szerkezetét láthatjuk.

28
5. ábra. A matrilinek domén szerkezete (Wagener és mtsai 2005).
2.4.2. A család tagjainak jellemzése és filogenetikai eredete
A matrilin-1 (CMP), a vWFA-szerű domént tartalmazó család egyik legegyszerűbb
tagja, egy homotrimér glikoprotein, 52 000 Da alegységeket hordoz. 3, 9% szénhidrátot,
főképpen N-kapcsolt oligoszacharidokat tartalmaz (Paulsson és Heinegard 1981). A
porc különböző formáiban, nagy mennyiségben előforduló fehérje. A szem
alkotóelemeiben is kimutatták (Stirpe és mtsai 1989), ennek alapján egy nagyon
specifikus funkciója feltételezett. Elektronmikroszkópos felvételen egy tipikus
virágcsokorszerű szerkezetet mutat, ahol az egyedülálló vWFA domének nem nyitottak,
hanem egymással kölcsönhatásban van a már említett vWFA1 és a vWFA2. A domének

29
3 egymással párhuzamos láncú, diszulfid-híddal kapcsolt, α-helikális coiled-coilokba
rendeződnek.
A matrilin-2 a legnagyobb molekulasúlyú családtag, 104 300 Da méretű az egérben, N-
terminális oligoszacharidokat tartalmaz (Piecha és mtsai 1999). Elektronmikroszkópos
vizsgálatok igazolták, hogy a matrilin-1-hez hasonlóan itt is a két vWFA domén
egymáshoz kötődik. Jellemző, hogy szöveti kivonatból nyert fehérjekivonatban minden
esetben egy igen heterogén összetételű fehérje mutatható ki, valószínűleg a proteolitikus
hasítás következményeként. A humán matrilin-2 313 AS ORF régiója 87% -os egyezést
mutatott az egér matrilin-2-vel. A 87%-os egyezés az AS szekvenciában az egér és a
humán matrilin-2 között azt mutatja, hogy valószínűleg a matrilin-2 funkcionálisan egy
nagy jelentőséggel bíró ECM komponens lehet az emlős szövetekben és szervekben.
(Deák és mtsai 1997). Néhány tény igazolja, hogy a matrilin-2 valóban a CMP közeli
rokona: 1) a vWFA-szerű domének hasonlósága; 2) a vWFA-és EGF-szerű domének
azonos sorrendisége. A közös domén szerkezetből feltételezhető hogy valószínűleg a
matrilin-1 és a matrilin-2 ugyanannak a közös génnek a duplikációjából származik,
amely egy duplikált vWFA-t, egy egyedülálló EGF-et, és egy oligomerizációs domént
tartalmazott.
A matrilin-3 a legkisebb családtag, többnyire a matrilin-1-el koexpresszálódik (Wagener
és mtsai 1997, Klatt és mtsai 2000). MALDI-TOF tömegspektrometriával az egér
matrilin-3 tömegét 49300 Da-nak találták. A második vWFA domén hiánya miatt az
alegységek nincsenek kölcsönhatásban önmagukkal ezért az elektronmikroszkópban ezt
a fehérjét több kifeszített, flexibilis karral látjuk.
A matrilin-4 egérben a két vWFA modul között 4 EGF- szerű domént tartalmaz.
(Wagener és mtsai 1998a,b, Klatt és mtsai 2001). A rekombináns fehérje tömege 72900
Da, ami 7%-os poszttranszlációs modifikációra utal. Noha a matrilin-4 két vWFA
domént tartalmaz, elektronmikroszkópban nem mutatható ki nyilvánvaló kölcsönhatás a
domének között (Klatt és mtsai 2001).
Összességében véve tehát a matrilinek egy közös ősből származnak, amelyben a vWFA
domének már megduplázódtak, mert az összes vWFA1 domén közelebb áll egy másik
molekula vWFA1 doménjéhez, mint a vWFA2-es modulhoz. A feltételezhető hipotézis
az, hogy a primordiális matrilin gén duplikálódik és ebből ered a matrilin-1 és matrilin-
3 valamint a matrilin-2 és matrilin-4 közös őse. Továbbá, a második génduplikációt

30
követően, a matrilin-3-ban a matrilin-1 vWFA2 doménjével megegyező vWFA domén
kiesett (Deák és mtsai 1999).
2.4.3. A matrilinek oligomerizációja és kölcsönhatása más ECM komponensekkel
Ahogy azt már említettem, a matrilin család C-terminális régióján lévő α-helikális
coiled-coil domén teszi lehetővé a molekula alegységek virágcsokorszerű
oligomerizációját. A teljes hosszúságú molekulák elektronmikroszkópos vizsgálata
kimutatta, hogy a matrilin -1 és matrilin-4 homotrimereket (Hauser és Paulsson 1994,
Klatt és mtsai 2001), míg a matrilin-2 és-3 homotetramereket képez (Pan és Beck 1998,
Piecha és mtsai 1999, Klatt és mtsai 2000). Az oligomerizációt E. coli-ban termeltetett
rekombináns coild-coil doménekkel is tanulmányozták (Frank és mtsai 2002). A
biokémiai és biofizikai vizsgálatok nemcsak igazolták a teljes hosszúságú molekulák
által képzett eddig ismert homo- és heteromer oligomereket, de újabb további hét
rekombináns matrilin lánckombinációra derítettek fényt. Termodinamikai mérésekkel
alátámasztották a heterooligomerek nagyobb hőstabilitását. A különböző matrilin
láncok kombinációja igen változatos képet mutat (6. ábra) és expressziós szint által
szabályozott. A teljes hosszúságú matrilin-2 láncokkal szemben a rekombináns coiled-
coil ccMat2 domének csak trimereket képeznek. A coiled-coil domének ugyanazon a
helyen oligomerizálódnak, mint a teljes hosszúságú fehérjék azzal az előnnyel, hogy
rájuk nem jellemző a szupramolekuláris szerveződés. A coiled-coilok funkcionális
szerepe abban rejlik, hogy a multimerek képződéséhez vezető alegységek
oligomerizációjában vesznek részt. Az oligomerizáció eredményeként többértékűség jön
létre, és a funkcionális helyek helyileg koncentrálódnak (Deák és mtsai 1999). Továbbá
a heteromerek növelik a matrilinek funkcionális diverzitását. A homo-és
heterooligomerek képződése a vWFA domén ligandkötő aktivitására és affinitására is
hatással van. (Ugyanez a jelenség jobban érthető a trombospondin vagy a laminin
hasonló példáján, amelyekre jellemző, hogy heparinkötő affinitása a homo-és
heterotrimerek képzésével fokozódik).
A vWFA doméneken keresztül a matrilinek kölcsönhatnak önmagukkal és egymással,
ami szupramolekuláris szerkezetek képződéséhez vezet, amelyek az ECM struktúrák
stabilizálásában vesznek részt.

31
A teljes fehérjék elektronmikroszkópos felvételén kimutatható, hogy a matrilin-1és a
matrilin-4 homotrimereket (Hauser és Paulsson 1994, Klatt és mtsai 2001), míg a
matrilin-2 és -3 homotetramereket képez (Piecha és mtsai 1999, Klatt és mtsai 2000).
Matrilin-1 és matrilin-3 heterooligomereket izoláltak újszülött humán – és borjú porcból
(Wu és Eyre 1998, Klatt és mtsai 2000). Matrilin-1 és matrilin-2, matrilin-1 és matrilin-
3, matrilin-1 és matrilin-4 valamint matrilin-2 és matrilin-4 heterooligomereket
mutattak ki. Matrilin-2 és matrilin-3, matrilin-3 és matrilin-4 heterooligomerek nem
voltak kimutathatók (Frank és mtsai 2002).
Egyéb molekulákkal történő kölcsönhatását illetően a matrilin-1-t elsőként az
aggrekánhoz kötve együtt tisztították (Paulsson és Heinegard 1979, Hauser és mtsai
1996). Még nem ismert, hogy ebben a molekula komplexben egyedülállóan az aggrekán
és a matrilin-1 vesz részt vagy egyéb matrilinek is képesek proteoglikánokhoz kötődni.
Elképzelhető, hogy azok a család más tagjaihoz kötődnek. A matrilin-1 a II típusú
kollagén fibrillumokhoz egy meghatározott periodikussággal kötődik (Winterbottim és
mtsai 1992) és részt vesz a kollagén-független filamentózus hálózat kialakításában is.
Az aggrekánhoz való kötődése kovalens kereszt-kötéseket vesz igénybe, ami a kor
előrehaladásával fokozódik. Azt pontosan nem tudjuk, hogy a CMP-nek melyik
doménje vehet részt ebben a kölcsönhatásban, de nagy valószínűséggel a vWFA-szerű
domén az, amelyik szerepet kap ebben. Mindkét vWFA-szerű doménnek jelentős
szerepe van a hálózatképzésben. Ha kiesik a vWFA2 domén, nem képződik multimer
matrilin-1 fehérje, ha hiányzik valamelyik vWFA domén, akkor nem képződik
kollagén-független hálózat sem (Chen és mtsi 1999). Porcsejt-tenyészetekkel végzett
kísérletek igazolták a matrilin-1 jelenlétét pericelluláris kollagén-függő és kollagéntól–
független filamentumokban egyaránt (Chen és mtsai 1995). A matrilin-1, matrilin-3 és a
matrilin-4 köti a kollagénV összetételű mikrofibrillumokat az aggrekán core proteinhez
és a kollagén II típusú fibrillumokhoz a biglikánon vagy a dekorin alkotta
csoportosuláson keresztül (Wiberg és mtsai 2003). Matrilin-1 defficiens egerekben nem
volt kimutatható elváltozás a kollagén II, IX, X, XI, aggrekán és COMP
expressziójában. Ennek alapján feltételezhető, hogy a matrilin-1 expressziójának
változása nem befolyásolja szignifikánsan ezeknek a molekuláknak a biokémiáját.
Valószínűleg itt is a többi matrilin fehérje redundáns funkciója érvényesül, mivel a
fejlődő csontban minden matrilin fehérje expresszálódik.

32
Immunfluoreszcenciás kísérletekkel igazolták, hogy a matrilin-2 sejtkultúra
tenyészetben képes saját molekulákkal alkotott, sejten kívüli hálózat létrehozására.
Emellett más, kollagén és nem kollagén természetű extracelluláris molekulákhoz is tud
kötődni, tehát részt vesz több fehérjéből felépülő, heteromolekuláris fehérjehálózatok
létrehozásában is.
A matrilin-2 a legnagyobb mennyiségben előforduló szöveti kollagénhez, a kollagén I-
hez kötődik. Ebben a kapcsolatban a tropokollagén molekula két terminálisát részesíti
előnyben. Plasma rezonanciás módszerrel igazolták a matrilin-2 kölcsönhatását a
kollagén II, III, IV és V molekulákkal is (Piecha és mtsai 2002 a). A matrilin-2-t
gyakran BM-hoz és mikrofibrillumokhoz asszociáltan írták le. Így számos más
molekulát is kimutattak matrilin-2-vel köcsönhatásban: a fibronectin, a laminin-1 és a
nidogén kötődését igazolták csonkolt matrilin-2-höz (Piecha és mtsai 2002 b).
ELISA-n alapuló ligand esszével mutatták ki a matrilin-4 molekula kötődését a
kollagén I, II, XI, XII, XIV, XXII molekulákhoz. Az említett molekulákhoz képest
néhány nem-kollagén molekula nagyobb affinitással köti a matrilineket, különösen a
COMP-ra (cartilage oligomeric matrix protein) jellemző ez a jelenség (Mann és mtsai
2004). Abból eredendően, hogy a COMP és a decorin is kölcsönhat a kollagénnel, a
matrilinek a kollagén rostokhoz elképzelhetően egy szendvics részeként kötődnek, ahol
a többi résztvevő komponens közvetlenül és erős affinitással kötődik a kollagénhez.
6. ábra. A matrilinek oligomerizációja. Lehetséges homo-és heteromerek. Piros-
matrilin-1, fekete-matrilin-2, zöld-matrilin-3, kék-matrilin-4 (Frank és mtsai
2002).

33
2.4.4. A matrilinek expressziója különböző szövetekben és szervekben
A szöveti eloszlásuknak megfelelően a matrilineket két csoportra oszthatjuk. A matrilin-
1 és -3 a váz elemek jelentős alkotóeleme, de felnőtt egérben kimutatták a matrilin-1
gén expresszióját extraszkeletálisan is (Aszódi és mtsai 1996, Klatt és mtsai 2002). A
vázképződés során a matrilin-1 fehérje és mRNS akkumulálódik a humán, egér és csirke
porc növekedési zónájában (Aszódi és mtsai 1994). A matrilin-1 expressziója térben és
időben egyaránt szigorúan szabályozott. Matrilin-1 ellenes antitesttel a fehérje
detektálható az embrionális myocardiumban az embriogenesis 9, 5 napján. Ez egy
tranziens expresszió és valójában nincs élettani jelentősége. A leghosszabb ideig tartó
matrilin-1 és matrilin-3 expressziót az emrionális kötőszövetben mérték, a fejlődés 12, 5
napján. A vázfejlődés előrehaladásával a matrilin-1és a matrilin -3 expresszálódó
doménjei megegyeznek a kollagén II doménjeivel. A fejlődés későbbi szakaszában a
porc felszíni rétegeiben ezeknek a matrilineknek az expressziója már nem olyan
kifejezett, mint a porc mélyebb rétegeiben (Segat és mtsai 2000). A kalcifikáció után a
matrilin -1 és a matrilin-3 a csontba épül. Immunhisztokémiával a matrilin-1-t
kimutatták a korneában, a szklerában, a placentában, a lencse tokban és a lencse
epithéliumában (Tsonis és Goetnick 1988, Kabosava és mtsai 2007). In situ
hibridizációval kimutatták a humán embrió retinájában (Mundlos és Zabel 1994).
Tranziens matrilin -1 expresziót mutattak ki embrionális egér szívben (Segat és mtsai
2000). A matrilin-3 megtalálható a porc minden típusában és a csontban egyaránt
(Wagener és mtsai 1997, Klatt és mtsai 2000), szélesebb spektrumú szöveti expressziót
mutatva a matrilin-1-hez képest. A matrilin-3 szöveti expressziója azonban születés után
leáll. Nagy valószínűséggel ez a két családtag, a matrilin-1, -3- az élet korai
szakaszában expresszálódik jelentős mértében, de a fehérjék lassú turnovere miatt
szöveti szinten az élet későbbi szakaszában is detektálhatók. A porcsejtek mátrixában a
matrilin-3-t egy filamentózus, sejteket összekötő, kollagén-függő hálózatban és egy
kollagén-független pericelluláris hálózatban is kimutatták.
A matrilin-2- a matrilin- 4 képezi a másik alcsoportot. Ezek sokkal elterjedtebbek a
szövetekben. Northern blot vizsgálatokkal, a porcot kivéve, magas expressziójukat
detektálták az uterusban, a szívben, a koponyatetőben és az agyban, alacsonyabb

34
expresszió volt detektálható a vesében, a vázizomban és a bőrben. A matrilin-1-hez
hasonlóan a matrilin-2 is elsőként az egér embrió szívében detektálható, de a matrilin-2
a matrilin-1-hez képest később jelenik meg, a fejlődés 10, 5 napján és szintézise nem áll
le, hanem folyamatos. A fejlődés későbbi szakaszaiban számos kötőszöveti sejt,
simaizomsejtek és egyes hámsejtek expresszálják (Piecha és mtsai 1999). Ezek a sejtek
a matrilin-2-t az őket körülvevő pericelluláris mátrixba halmozzák fel. Jóllehet sok
esetekben BM-al asszociált, még ezidáig sem tisztázott integráns BM fehérje volta.
Más esetekben a matrilin-2 egy filamentumokból felépülő hálózat komponense. Jelen
van a porcban is. Minden eddig vizsgált szervben a matrilin-2 fehérje exszpresszió a
mért mRNS expresszióhoz képest alacsonyabb szinten volt. Az egér matrilin-2 gén
expressziót detektálták laza és tömött kötőszövetekben, izomsejtekben és néhány
epitheliális sejttípusban (Deák és mtsai 1997, Piecha és mtsai 1999), szubepitheliális
kötőszövetekben, bőrben és emésztő traktusban, vérerek falában. Matrilin-2 mRNS-t
detektáltak a szív kötőszöveti elemeiben, simaizomsejtekben, méhizomzatben és az
agyban (Deák és mtsai 1997, Sharma és mtsai 2006).
Az összes matrilin közül a matrilin-4 a legáltalánossaban előforduló, és mindenhol
kimutatható, ahol a többi matrilin megjelenik (Klatt és mtsai 2001). A matrilin-2-höz
hasonló a matrilin-4 szöveti expressziója: tömött- és laza- rostos kötőszövetben,
csontban, porcban, idegrendszerben és számos más szervben fordul elő (Wagener és
mtsai 1998a; 1998b). Részleges átfedést mutat a matrilin-2-vel. Az idegrendszeri
kifejeződése sokkal kifejezettebb, mint más matrilineké, így például az agy a
leggazdagabb matrilin-4-ben (Klatt és mtsai 2001).
A szöveti kivonatokban a matrilin-2 és matrilin-4 általában degradált, fragmentálódott.
Továbbá, a teljes hosszúságú fehérjékhez képest a fragmentekből hiányzik egy vagy
több alegység (Klatt és mtsai 2001). A matrilin-4 rekombináns formáját humán
embrionális vese - 293 sejtekben expresszálva, monomer, dimer és trimer fragmentek
keverékét kapták (Klatt és mtsai 2001).
2.4.5. A matrilinek szerepe betegségekben
A matrilin fehérje családban a matrilin-3 az egyetlen olyan családtag, amelynek
mutációja az eddigi ismeretek alapján a chondrodysplasia különböző formáival van
kapcsolatban (Aszódi 1998, 1999, Leighton és mtsai 2007).

35
A matrilin-1 és matrilin-3 mutánsok fenotípusosan normális vázrendszerrel
rendelkeznek, de biokémiai és ultrastruktúrális vizsgálatokkal eltéréseket lehetett
kimutatni. Ezt matrilin-1, -2 és -3 knock-out egereken is igazolták. Mind a matrilin-1,
mind a matrilin-3 modulálja a kollagén fibrillogenezisét a porcban, a normálisan
megjelenő fenotípus a matrilinek egymással történő kompenzációjára utal. A matrilin-
1/-3 kettős knock-outokban átmenetileg megemelkedő matrilin-4 homotrimer is a
kompenzációs mechanizmust támogatja, míg más elméletek szerint a funkcionális
redundancia lehetősége merül fel a matrilin molekulák és a kölcsönható molekulák
között (COMP, dekorin, biglikán). A COMP, matrilin-3 és kollagén IV károsodását
írták le pseudochondro-dysplasiában (PSACH). A multiplex epiphysealis dysplasia
domináns formájában (MED) (Briggs 2004), és az örökletes bilaterális
microepiphysealis dysplasiában (BHMED) is detektálható a matrilin-3 mutációja. A
mutáció elsősorban a vWFA domén β-lemezét érinti (Chapman és mtsai 2001, Jackson
és mtsai 2004, Mabuchi és mtsai 2004). A matrilin-3-ban a mutáció az első EGF-szerű
doménben is lehet, így például osteoarthritisben az EGF-ek első doménjében
bekövetkező cystein-serin csere okozza az S-S diszulfid-híd képződésében létrejövő
károsodást (Stefánsson és mtsai 2003). MED-ben nem írtak le matrilin-1 mutációt.
Ezzel szemben német populáció osteoarthritises betegeiben matrilin-1 gén mutációt,
matrilin-1 és matrilin-3 overexpressziót találtak (Okimura és mtsai 1997). Összefüggést
írtak le a felnőttkori idiopatikus szklerózis (IS) és a matrilin-1 expressziója között. Egy
igen ritka autoimmun porcbetegségben, amely elsősorban a külső fület és a tracheát
érinti a visszatérő polychondritisben (PA) matrilin-1 ellenes immunválaszt mutattak ki
(Buckner és mtsai 2000), rheumatoid arthritisben szenvedő betegekben emelkedett
volt a matrilin-1 fehérje expresszió szintje (Saxne és mtsai 1989).
Matrilin-2 és a matrilin-4 mutációval összefüggő genetikai elváltozásokat nem sikerült
kimutatni. Matrilin mutáns egerekben nem volt kimutatható fenotípusos elváltozás,
noha megfigyelhető volt az abnormális kollagén fibrillumok felhalmozódása és a
kollagén molekulák átmérőjének növekedése (Aszódi és mtsai 1999, Ko és mtsai 2004,
Mátés és mtsai 2004). A matrilin-2 hiányos egerek látszólagos normális fejlődése
többféleképpen is értelmezhető. Egyik kézenfekvő magyarázat az, hogy más matrilin,
vagy a sejtközöttti állomány más fehérjéje átveheti azokat a szerepeket, amelyeket a
matrilin-2 betölt. Erre azért gondolhatunk, mert a matrilinek szerkezeti felépítése és

36
szöveti előfordulása igen hasonló, részlegesen átfedő, továbbá nemcsak homo-, hanem
hétféle heterooligomert is tudnak formálni. A funkcionális redundancia lehet az oka
annak is, hogy eddig nem írtak le olyan súlyos fejlődési rendellenességet, amit a
matrilin gének mutációja idéz elő. Egérben indukált fibrotikus vese modellben (TIF =
renal tubulointerstitial fibrosis) az ECM-re fókuszált klaszter analízissel, a betegséggel
asszociált gének között a matrilin-2 down-reguláltságát mutatták ki (Sadlier és mtsai
2004). Összességében véve elmondható, hogy a matrilinek in vivo betöltött funkciója
még máig sem egészen pontosan tisztázott. Szupramolekuláris struktúrákban betöltött
„adaptor” szerepük ismert, noha ennek részletes tanulmányozását e molekulák
redundanciája nehézkéssé teszi. A sejt-mátrix kölcsönhatásokban is szerepük lehet: a
sejt felszíni filamentumok között foglalnak helyet, de azonkívül, hogy integrin
receptorok, még nem sokat tudunk a sej-sejt illetve sejt-mátrix kölcsönhatásban betöltött
szerepükről.
2.4.6. A matrilin-2 expressziója humán szövetekben és tumorokban
Humán szerveket illetően kevés tudásunk van a matrilin-2 expressziójáról. Bőrben és a
bőr függelékekben, az erek BM-nak legalsó rétegében mutattak ki matrilin-2
expressziót. In situ hibridizációval a keratinocytákban és a fibroblastokban is sikerült
mRNS expressziót detektálni. Noha mindkét sejttípus expresszálta a matrilin-2-t,
különböző módon történik a fehérje processzálása. A fibroblastok, úgy tűnik, leginkább
fokozzák a matrilin-2 expresszióját (Piecha és mtsai 2002 b). A keratinocyták termelik,
majd a szubepidermális szövetek felé transzportálják, a kötőszöveti stroma irányában.
Ez a mechanizmus egészen pontosan nem ismert, de feltehetőleg specifikusan
kölcsönható partner molekulák bizosítják a matrilin-2 BM-ban történő lokalizációját
(Piecha és mtsai 2002 a, b).
Az egyik leggyakoribb gyerekkori primer agytumorban, a pilocitikus astrocytomában
(PA) és multiform glioblastoma (GBM) mintákban mérték az mRNS szintjét real-time
RT- PCR módszerrel. A PA tumorokban a matrilin-2 mRNS szintje hétszer olyan magas
volt, mint a tumor által nem érintett agyi fehérállományban. A GBM minták nem
mutattak matrilin-2 fehérjeszint emelkedést. A magas matrilin-2 szint tehát
összefügghet klinikailag agresszívebb tumorok kialakulásával és hozzájárulhat a tumor

37
terjedéséhez. A matrilin-2 overexpresszió prognosztikus marker lehet PA tumorokban
(Sharma és mtsai 2006).
A matrilin-2-ről májra vonatkozó adatokat mostanáig még nem találtunk az
irodalomban. Minthogy ECM és BM komponens, így jelentősége felmerülhet mind a
májregenerációban mind, pedig a máj cirrhotikus és tumoros elváltozásaiban.
A ECM lehetséges célpontot jelenthet a daganat terápiában. A közelmúltban
beigazolódott, hogy az ECM mikrokörnyezet jelentősen módosítani képes az egészséges
és a malignus sejtek fenotípusát, a tumorsejtek szaporodását, invaziv és metasztatikus
képességét, valamint daganatellenes szerekkel szembeni válaszreakcióját (Hodkinson és
mtsai 2007). Ez utóbbi is kiemeli e nemrégiben leírt mátrix molekulák vizsgálatának a
jelentőségét a HCC-ben és cirrhosisban egyaránt.

38
3. CÉLKÍTŰZÉSEK
Az előzőekben vázolt irodalmi áttekintés alapján vizsgálataink a következő kérdések
megválaszolását célozták meg:
1. Kimutatható-e matrilin-2 mRNS és fehérje expresszió normális patkány és
humán májban?
2. Milyen mértékben és hol expresszálódik a matrilin-2 a májregeneráció
során?
3. Milyen jelentősége lehet a matrilin-2-nek a májregenerációban?
4. Milyen sejteknek lehet szerepe a matrilin-2 termelésében a májregeneráció
során?
5. Hogyan változik mennyiségében és lokalizációjában a matrilin-2
expressziója a cirrhotikus májban, összehasonlítva normál májszövettel?
6. Van-e eltérés a matrilin-2 expressziójában a tumoros májszövetben a normál
májakhoz képest?
7. Milyen különbség figyelhető meg a matrilin-2 expresszió szempontjából az
eltérő eredetű (cirrhotikus és nem cirrhotikus talajon létrejött)
hepatocelluláris carcinomákban?
8. Van-e eltérés a matrilin-2 expressziójában a különböző differenciáltságú
tumorokban?

39
4. ANYAG ÉS MÓDSZER
4.1. A vizsgálatokhoz használt oldatok, pufferek összetétele
Oldat neve Összetétel DEPC víz DEPC (Sigma D 5758), 1l deszt víz
10xPBS 1,5 M NaCl (Sigma, S9625), 12,2 Na2HPO4 (Sigma, S0876), 0,014mM
K2HPO4 (Interkémia, 04190203)
10x TBE 1 M Tris (GIBCO, 15504-020), 0,83 Bórsav (GIBCO, 15705-023), 0,01 M
EDTA(GIBCO 15576-028)
10x Tris-HCl 0,05 M Tris (GIBCO, 15504-020), 0,3 M NaCl (Sigma, S9625)
TBS 0,05 M Tris (GIBCO, 15504-020), 0,15 M NaCl (Sigma, S9615)
TBS-Tween 0,05 M Tris (GIBCO, 15504-020), 0,15 M NaCl (Sigma, S9615), 0,1%
Tween
20x SSC 3 M NaCl (Sigma, S9625), 0,3 M Na –citrát (Sigma, S 4641)
10%-os
Pufferolt
formalin
40% Formalin (Sigma, 8775), 6,5g Na2HPO4 (Sigma, S 0876), NaH2PO4
(Sigma, S0751)

40
4.2. Állatkísérletek
4.2.1. Parciális hepatectomia (PH)
A beavatkozás 180-200g súlyú hím Fisher 344 (F- 344) patkányokon történt Higgins és
Anderson klasszikus módszere szerint (Higgins és Anderson 1931, Grisham és mtsai1997) az
SE I. sz Patológia és Kísérletes Rákkutató Intézetében. A vizsgálatokat a parciális
hepatectomiát követően 24, 48, és 72 órával a műtét után vett szövemintákon végeztük. Az
altatás dietil-éterrel történt. A májminták egy részét formalinban fixáltuk, más részét
folyékony nitrogénben fagyasztottuk további vizsgálatok céljából.
4.2.2. Parciális hepatectomia/acetylaminofluorén (PH/AAF) modell
Az állatok az 1 %-os dimethylcellulózban 2mg/ml koncentrációban oldott
acetylaminofluorént (AAF) 5 mg/kg dózisban kapták hat egymást követő napon át. Ezt
követően parciális hepatectomia történt a 7. napon, amit ezután még hat AAF-es kezelés
követett. Az állatok leölése a kezelés során különböző időpontokban történt (a PH-t követő 9.,
11. és 13. napokon) az ovális sejt proliferáció csúcspontjának meghatározása végett.
Az állatkísérleteket a Nemzetközi Állategészségügyi Intézet kísérleti protokolljaival
megegyező módon végeztük el.
4.3. Humán szövetek
A Semmelweis Egyetem II. sz. Patológiai Intézetében archivált valamint újonnan gyűjtött
humán májmintákat használtuk fel vizsgálatainkhoz, amelyeket formalinban, illetve RNA-
laterben (Sigma) fixáltunk vagy folyékony nitrogénben történt fagyasztás után -80◦C-on
tároltunk. A sebészileg rezekált anyagok az SE I. sz. Sebészeti Klinikáról, a kontrollként
használt egészséges normál májak pedig balesetben elhunyt személyekből származtak, az SE
Igazságügyi Orvostani Intézetéből. A vizsgálatokhoz TUKEB engedéllyel rendelkezünk (#
172/2003, # 2/2004).
A vizsgált HCC-k csoportosítása az Edmonson-Steiner féle klasszifikáció alapján történt (G1-
G3) (1. táblázat). A nemek szerinti megoszlás a következő volt: 27 férfi, 8 nő. Az átlag
életkor 65 év (44-82). A 35 eset közül 15 esetben a hepatocelluláris carcinoma cirrhosis
talaján fejlődött ki. A többi esetben enyhe és igen mérsékelt mértékű fibrosis volt jellemző a

41
tumort környező májszövetre, de sem nodulus képződés, sem pedig a cirrhosisra jellemző
duktuláris reakció nem volt megfigyelhető. A 15 eset közül 7 minta volt HCV-vel fertőzőtt.
1. táblázat. A vizsgálatokhoz használt humán májminták.
Vizsgált májminták esetszám
Normál májminták 10
HCC környező tumormentes máj 35
HCC 35
I.
1. cirrhotikus talajú HCC 15
2. nem cirrhotikus talajú HCC 20
II.
1. jól differenciált HCC (G1) 19
2. közepesen differenciált HCC (G2) 9
3. rosszul differenciált HCC (G3) 7
4.4. Morfológiai vizsgálatok
4.4.1. Hisztológia
A patkány máj, valamint a sebészetileg eltávolított tumor mintákat a környező nem tumoros
szövetekkel együtt, valamint a normál humán máj mintákat 10%-os neutrálisan pufferolt
formalinban fixáltuk 24 órán keresztül, utána dehidráltuk és paraffinba beágyaztuk. A
morfológiai elváltozások kimutatása és a tumor grade megítélése 3-4 µm-os hematoxilin-
eozinnal (HE) festett metszeteken történt. Kiegészítő PAS, Shikata-féle orcein, Picrosirius-
vörös festéseket rutinszerűen elvégeztük.
4.4.2. Immunhisztokémiai vizsgálatok
4.4.2.1. Paraffinos metszeteken végzett immunfestés A formalinban fixált, paraffinba ágyazott mintákból 3-4 µm-es metszeteket készítettünk, majd
deparaffináltuk: kétszer 10 percig xilolos, majd kétszer 5 percig etanolos áztatással. Desztillált
vízzel történő mosás után az endogén peroxidáz blokkolást 3%-os H2O2-ban végeztünk 20
percig. PBS pufferes mosás után az antigénfeltárás proteáz emésztéssel történt (ready-to-use

42
proteináz-K, 10 perc, 25◦ C, DAKO, CA, USA). Újabb PBS-sel történő mosás után az antitest
aspecifikus kötődéseket 1%-os normál ló szérummal illetve 3%-os BSA-val (PBS) blokkoltuk
20 percen át szobahőmérsékleten. Ezután a megfelelő elsődeges antitest oldatával egy
éjszakán át 4 ◦C-on inkubáltunk. PBS-ben történt mosásokat követően 20 percig a másodlagos
antitesttel inkubáltuk a metszeteket. A biotinilált másodlagos antitestnél ezt az ABC inkubálás
követte.
A matrilin-2 kimutatása végett az immunreakció érzékenységét egy nagy érzékenységű
jelamplifikációs immuntechnika alkalmazásával, biotinilált tiraminnal fokoztuk (ABC-BT
módszer) (7. ábra) Az immunreakció Merz és munkatársai által leírt módszer szerint történt
(Merz és mtsai 1995). Az ABC inkubálást biotinilált tiraminos inkubálás követte (7mM
biotinált tiramin törzsoldatból 1: 125 + 0,033% H2O2 (PBS-ben) 10 perc, 37 ◦C). Ismételt
ABC reakció után az immunreakciót 3-amino-9-ethyl-carbazol (AEC) (Biogenex HK 129-5K,
San Ramon, USA) vagy 3,3’-diaminobenzidin tetrahidroklorid (DAB) (Genetex, San Antonio,
USA) kromogénekkel hívtuk elő. A háttérfestés Mayer-féle hematoxilinnal történt és
glicerines fedőanyaggal fedtük a metszeteket. A kontroll metszeteknél az elsődleges antitesttel
való inkubálást elhagytuk.
4.4.2.2 Fagyasztott metszeteken végzett immunfloureszcenciás vizsgálat 6 µm vastag fagyasztott metszeteket készítettünk kriosztáttal (SHANDON CC 0053000603,
Patosystems, Minnesota, USA), ezt követően 10 percig metanolban fixáltuk – 20◦C-on. A
fixálás után a metszeteket TRITON tartalmú PBS-ben (pH 7,4) tártuk fel 5 percig. A
metszeteket 1 éjszakán át 4 ◦C-on inkubáltuk az elsődleges antitesttel. Ezt követően egymás
után háromszor 5 percig mostuk a metszeteket PBS-ben. A reakciót Alexa Flour-488-
konjugált- anti-egér IgG másodlagos antitesttel tettük láthatóvá (Molecular Probes, Oregon,
USA). A metszeteket DAPI-t tartalmazó Vectashield Mounting Mediummal fedtük le (Vector
Laboratories, Burlingame, CA, USA). Az immunhisztokémiai vizsgálatokhoz használt
antitesteket a 2. táblázat foglalja össze.
4.4.2.3. Fotódokumentáció A fotókat Olympus BX mikroszkóphoz (Tokyo, Japán) rögzített kamera segítségével
készítettük. Az immunfluoreszcenciás képeket konfokális lézer-szcanning mikroszkóppal
elemeztük és a hozzá tartozó szoftverrel fotóztuk (Bio-Rad MRC-1024 system, Bio-Rad,
Richmond, CA, USA).

43
2. táblázat. Az immunhisztokémiai vizsgálatok során használt antitestek
Elsődleges antitestek
• poliklonális nyúl anti - matrilin-2 (1:150 hígításban (PBS), ajándék dr Deák Ferenc
munkacsoportjától, Szeged, SZBK, Piecha és mtsai 1999)
• monoklonális egér anti-laminin (1:100 hígításban (PBS), Novocastra, Newcastle, UK)
• poliklonális nyúl anti-laminin (1:100 hígításban (PBS), DAKO)
• monoklonális anti-egér OV-6 (1:100 hígításban (PBS), R&D, Minneapolis, USA)
• monoklonális CD34 (1:100 hígításban ( PBS), DAKO, clone QBEnd/10)
Másodlagos antitestek
• biotinilált anti-nyúl IgG (ABC kit, Novocastra, Newcastle, UK)
• biotinilált anti-egér IgG (ABC kit, Novocastra, Newcastle, UK)
• Alexa Flour-488-konjugált anti-egér IgG (1: 100 hígításban (PBS), Molecular Probes,
Oregon, USA)
• Alexa Flour-568-konjugált anti-nyúl IgG (1: 100 hígításban (PBS), Molecular Probes,
Oregon, USA)

44
7. ábra. Az ABC –BT: biotinilált tiramin jelamplifikációs módszert bemutató sematikus
ábra (Füle és Kovalszky 2002)
A mintában található jelet (a kimutatni kívánt antigént) specifikus ellenanyaggal reagáltatjuk.
Ehhez kötjük a biotinnal jelzett másodlagos ellenanyagot. A következő lépésben avidin.-
biotin - peroxidáz komplexet adunk a mintához, ekkor az avidin a biotinhoz kötődik. A
peroxidáz a következő lépésben alkalmazott hidrogénperoxidot redukálja, a szabad gyökként
viselkedő nascens oxigén felszabadulási helyére bekötődik a biotinált tiramin, ami az ABC-
vel újra amplifikálható. A reakciót végül AEC-vel, a hidrogén-peroxidáz szubsztrátjával
tesszük láthatóvá. A tiraminhoz más molekulát kötve (fluoreszcein, digoxigenin) a reakció
fluoroszcens kimutatásra is használható.

45
4.4.2.4. A matrilin-2 immunhisztokémiai reakciók morfometriai elemzése A matrilin-2 immunhisztokémiai reakciókat fénymikroszkóp segítségével vizsgáltuk
(200X nagyítás, Olympus BX). Tíz egymást nem fedő random kiválaszott területet
értékeltünk ki. A matrilin-2 immunreakciót rögzítő digitális képeknek az
immunreakciók intenzitására és a pozitív területek meghatározására a Leica QWin
programot (Leica Microsystems Imaging Solutions Ltd., Cambridge, UK) használtuk
fel. Az immunpozitív területet kijelölve végeztük el a méréseket. A vörös, zöld, kék
összetevők alapján állítottuk be a küszöbértéket, amelyet minden metszet esetében
állandónak fogadtunk el az összevethetőség érdekében. Az értékelésre kiválasztott
területen azokat a reakciókat fogadtuk el pozitívnak, amelyek a küszöbértéket
meghaladták.
4.5. Molekuláris biológiai vizsgálatok
4.5.1. Szöveti lézer mikrodisszekció és RNS izolálás patkány májból
AAF/PH kezelt és a kontroll normál májakból 10 µm vastag fagyasztott metszeteket
készítettünk, acetonban fixáltuk és szobahőmérsékleten szárítottuk majd RN-áz mentes
HE-al festettük. A kiválasztott területeket lézer katapult technikával vágtuk ki a
metszetekből RNS izolálás és PCR vizsgálat céljából. A célterületek kivágásánál
nagyfokú óvatossággal jártunk el az RN-áz mentes környezet megőrzése miatt. A
mikrodisszekciót a PALM lézer mikrodisszekciós rendszer segítségével végeztük a II.
Belgyógyászati Klinika kutatólaboratóriumában. A kivágott sejteket steril Eppendorf
csövek kupakjában gyűjtöttük, 500-1000 ovális sejtet és hepatocitát gyűjtöttünk össze (a
PH-t követő 13. napon az AAF/PH-kezelt állatokból) lysis pufferbe (Qiagen). Az RNS
izolálást RNeasy Mini Kit-tel (Qiagen) végeztük el, a gyártó utasítása szerint.
4.5.2. RNS-izolálás RNA laterben fixált humán anyagból
13 HCC-t, ugyanennyi környező mintát és 10 normál májat használtunk fel RNS
izoláláshoz. Egyenként kb. 20 mg RNA laterben (Sigma) fixált szövetből Trizol
(Invitrogen, Carlsbad, CA, USA) módszerrel RNS-t izoláltunk (Mannhalter és mtsai

46
2000). Homogenizálás után a szövethomogenizátumot Trizol reagensben inkubáltuk
szobahőmérsékleten 15 percig, majd 0,2 ml kloroform hozzáadása után vortexeltük és
centrifugáltuk. Ezután a felülúszót, amely az RNS-t tartalmazta, kloroformmal tovább
tisztítottuk. Az RNS-t izopropanollal kicsaptuk, vortexeltük és tíz perc állás után újra
centrifugáltuk. Ezután leöntöttük a felülúszót és 75 %-os etanollal mostuk, majd
centrifugáltuk. Roche RNS tisztító kitből (Roche, 3270289, Indianapolis, USA)
származó elúciós pufferben oldottuk az RNS-t, DNázzal emésztettük és tisztítottuk. A
kapott pelletnek spektrofotométerrel megmértük az optikai denzitását (O.D.) és az RNS
koncentrációját. További felhasználásig a tisztított RNS-t -80°C-on tároltuk. Az RNS-ek
integritását 1%-os agaróz gélen történt futtatással ellenőriztük.
4.5.3. cDNS előállítás
1 µg-nyi mennyiségű totál RNS felhasználásával egy RT-PCR reakcióban cDNS-t
állítottunk elő. A reakcióhoz random hexamert és M-Mulv reverz transzkriptázt
használtunk. A reakcióelegy összetételét a 3. táblázat foglalja össze.
3. táblázat. A cDNS szintézis komponensei
Komponensek
Térfogat
A kompnenst gyártó cég
RT Puffer 2 µl Applied Biosystems, CA, USA
Hexamer 0,5 µl Applied Biosystems, CA, USA
dNTP mix 0,5 µl Roche D 8220, Indianapolis, USA
20 U/ µl RN-áz gátló 0,5 µl Applied Biosystems, CA, USA
50U/ µl Reverz Transzkriptáz 1 µl Applied Biosystems, CA, USA
dH2O 4,5 µl Eppendorf 0032006159
RNS templát 1 µg
Összesen 10 µl
A reakció paraméterei:
25°C 10 perc
42°C 50 perc
95°C 5 perc,
4°C ∞

47
4.5.4. Primerek és a PCR körülményei
A kísérletek megkezdése előtt a rendelkezésre álló szekvenciák alapján a Primer
Expressz 2 program (Applied Biosystems, CA, USA) segítségével megterveztük a
lehetőségek szerinti legideálisabb primereket. A tervezésnél figyelembe vettük, hogy
bizonyos fehérjéknek több transzkripciós variánsa is létezik; ezeknél olyan primereket
használtunk, amely valamennyi variáns expressziójának kimutatására alkalmas. A
tervezés eredményei az 4. táblázatban összefoglalt primerek:
4. táblázat. A kísérletek során alkalmazott primerek.
Primer neve Orientáció Szekvencia (5’-3)
HUMÁN PRIMEREK
MATN2 forward GACGGAAGACGGTGCAAGAA
MATN2 reverse CCAGTGACAAACTGCTTCACGA
GAPDH forward CATGGGTGTGAACCATGAGAAGT
GAPDH reverse TGGACTGTGGTCATGAGTCCTT
PATKÁNY PRIMEREK
Matn2 forward AAACTGGACTCTTGTGCTTTGGG
Matn2 reverse AAACTGGACTCTTGTGCTTTGGG
Gapdh(rat) forward TGGAGAAACCTGCCAAGTGTG
Gapdh(rat) reverse GAGTGGGAGCTGCTGTTGAAGT
4.5.5. Grádiens PCR
A PCR-hez beállítottuk az optimális primer kötődési hőmérsékletet grádiens PCR-rel
(Merck, Eppendorf Mastercycler Gradient). A reakcióhoz használt komponenseket az 5.
táblázat tartalmazza. Az azonos tartalmú PCR csöveket úgy helyeztük el a készülékben,
hogy 53°C és 66°C közötti hőmérsékleten történjen meg a primer kötődés.

48
5. táblázat. A grádiens PCR-hez használt komponensek.
Komponensek
Térfogat
A komponenst gyártó cég
10x PCR puffer MgCl2dal 2,5 µl Roche J 0051
10mM dNTP mix 0,5 µl Applied Biosystems, D8220
25 µM forward primer (MATN2) 0,5 µl
25 µM forward primer (MATN2) 0,5 µl
Csertex
Csertex
5U/ µl AmpliTaq Gold Enzim 0,5 µl Roche G0448
dH2O 19,5 µl Eppendorf 0032006159
cDNS 1 µl
Összesen 25 µl
A reakció paraméterei:
kezdeti denaturálás 95°C 10 perc
denaturálás 95°C.............. 30 másodperc
primer kötődés 53-66°C ....... 30 másodperc
szintézis 72°C ............. 30 másodperc
leállítás 4°C………… ∞
Ciklusszám: 35
A következő hőmérsékleteken vizsgáltuk a primerek kötődését: 53, 2°C, 55, 5◦C, 58,
1°C, 60, 8°C, 63, 5°C, 66°C.
4. 5. 6. Agaróz gélelektroforézis
A grádiens PCR reakciók során nyert termékeket 2%-os agaróz gélben futtattuk meg (8.
ábra). A gélkészítésnél 1 g agarózhoz (GIBCO 15510-019) 50 ml 1x-es TBE puffert
adtunk, majd forraltuk és 2, 5 µl etidium-bromidot (BIO-RAD 161 0433) adtunk hozzá.
A mintákat (12 µl) mintafelvivő pufferben (Novagen 69180) futtattuk párhuzamosan a
DNS létrával (5 µl) (Novagen 70539), 1x TBE pufferben. A futtatás 100 V-on 40 mA
áramerősség mellett 20-30 percig történt. Ezután a SYNGENE MultiGenius Bio
Imaging rendszerével GeneSnap szoftver felhasználásával lefotóztuk a gélt. Ez alapján a
primer legoptimálisabb kötődési hőmérsékletének a 63°C találtuk. A későbbiekben

49
elvégzett real-time RT-PCR reakciókban ezt állítottuk be primer kötődési
hőmérsékletként. A továbbiakban ezen a hőmérsékleten végeztük a reakciókat.
4.5.7. Real-time RT-PCR
A real-time RT-PCR-hez mintánként 12, 5 µl Sybr Green Master Mixet (BIO-RAD
1708882), illetve az általunk megtervezett primerekből 500 nM koncentrációban 0,5-0,5
µl-t és a korábban előállított cDNS-ből 2 µl-t, valamint 9,5 µl desztillált vizet
használtuk fel. A reakció paraméterei:
Kezdeti denaturálás.............95°C .........2 perc
Denaturálás .........................95°C .........20 másodperc
Primer kötődés ....................63°C .........30 másodperc
Szintézis..............................72°C .........30 másodperc
Ciklusszám: 40
A termék homogenitásának ellenőrzése miatt olvadáspont analízist végeztünk el 55 °C -
95°C-on (9. ábra), mely a termékek homogenitását és a primer specifikus kötődését is
alátámasztotta. A mérések 96-os plate-eken, duplikátumokban történtek a kontroll
génekkel párhuzamosan futtatott reakcióban. A PCR termékek méretének ellenőrzésére
a reakció termékekből 10 µl-t futtattunk 2%-os agaróz gélen, majd etídium-bromidos
festéssel tettük láthatóvá az amplikonokat (9. ábra).
1 2 3 4 5 6 L
8. ábra. Matrilin-2 grádiens PCR termékek
(1 - 53,2 ◦C; 2 - 55, 5◦C; 3 - 58,1◦C; 4 - 60,8 ◦C; 5 - 63,5 ◦C;L - DNS létra)
100 bp

50
9. ábra. A. Matrilin-2 real-time RT-PCR termékek agaróz gélen. B. A termékek
olvadáspont analízise.
4.5.8. Real-time RT-PCR eredmények elemzésének módszere
A kapott adatokat a REST (Relative Expression Software Tool) néven ismert relatív
kvantifikálásra alkalmas számítógépes program segítségével értékeltük ki. (Pfaffl és
mtsai 2002). A vizsgálatokhoz a GAPDH háztartási gént használtuk, ezekhez
viszonyítottuk az általunk vizsgált gének expresszióját, az alábbi képletet figyelembe
véve.
(E célgén) ▲CPcélgén (Ct kontroll – Ct minta)
(E referencia gén) ▲CPreferencia gén (Ct kontroll – Ct minta)
R= a relatív expresszió;
E = a PCR reakció hatékonysága (azaz a ciklusonként észlelt PCR növekedés szorzója);
CP = crossing point, az a ciklusszám, ahol az RNS mennyisége meghaladja a
detektálhatóság küszöb (treshold) értékét;
t - treshold, küszöbérték: exponenciális fázisban;
Ct – treshold ciklus: hányadik ciklusban lépi át az adott minta fluoreszcenciája ezt a
szintet – a kiindulási DNS mennyiséggel arányos;
A
R =
B

51
Ct kontroll – a kontrollnak vett minta treshold ciklus értéke, Ct minta- a minta treshold
ciklus értéke.
A statisztikai kiértékelést mind az immunhosztokémiai reakciók esetében, mind pedig a
real-time RT-PCR eredményeknél a GraphPad Prism 2.01 (GraphPad Software, Inc.
San Diego, CA, USA) program segítségével végeztük. A csoportok közötti
összehasonlításra a Mann-Whitney U tesztet alkalmaztuk. A különbséget p < 0,05
esetén vettük szignifikánsnak.
4.5.9. HCV vírus detektálása humán májmintákból Nested - PCR módszerrel
A módszer két egymást követő PCR lépésből áll, melynek során az első amplifikálás
alatt felszaporított nagyobb DNS fragmentről a második lépésben egy kisebb
bennfoglalt szakaszt amplifikálunk. A módszer növeli a DNS amplifikáció
érzékenységét azáltal, hogy egy nem specifikus DNS amplifikáció is történik a reakció
során. Ez azt jelenti, hogy a HCV esetében egy olyan – általánosabb DNS szakaszt
felszaporítunk, amely a vírus több típusában is megtalálható. Ehhez két primer párt
használunk. Az első amplifikálás során nyert DNS terméket használjuk a második
PCR-ben templátként. A második lépésben a felszaporított fragmentről amplifikáljuk a
belső, specifikus primerpárok segítségével az adott HCV típusra egyedien jellemző
DNS szakaszt (10. ábra).

52
10. ábra. A Nested PCR elvét bemutató sematikus ábra.
Az első lépésben a cDNS szintézist a külső primer pár forward primerével (HCV-8)
végeztük el. A használt reakcióelegy és paraméterek a 6. táblázatban vannak feltüntetve.
6. táblázat. HCV- cDNS szintézishez használt komponensek
Komponensek
Térfogat (µl)
A komponenst gyártó cég
RT puffer 5x 2 Boehringer 1465376
HCV-8 primer 0,5 Genset 351330F
10 mM dNTP mix 0,5 Roche D 8220
20 U/ µl RN-áz gátló 0,5 Applied Biosystems, D8220
50 U/ µl Reverz Transzkriptáz 0,5 Boehringer 84106920
H2O 5 Eppendorf 0032006159
DNS 1
Összesen 10
Külső primer pár Külső primer pár Target DNS
Belső primer pár Belső primer pár
Az első amplicon
A target DNS specifikus amplifikációs terméke

53
A reakció paraméterei:
25◦C 10 perc, 42◦C 30 perc, 48◦C 10 perc, 95 ◦C 5 perc, 4◦C – on leállítás.
A második körben a kapott cDNS-t amplifikáltuk fel a cDNS előállítás során használt
reakcióelegyében fölöslegben maradt forward primerrel és a külső reverse (HCV-11)
primerrel. A reakcióelegy összetételét a 7. táblázat tartalmazza.
7. táblázat. Az első amplifikációs körben használt reakcióelegy összetétele.
Komponensek
Térfogat µl
A komponenst gyártó cég
10x PCR puffer 5 Roche J0051
Taq Gold enzim 0,5 Roche G0448
10 mM dNTP mix 1 Roche D 8220
HCV-11 primer 1 Genset 351332
dH2O 38,3 Eppendorf 0032006159
RNS templát 5
Összesen 50 µl
Reakció paraméterek: kezdeti denaturáció 95◦ C 10 perc
denaturáció 95◦ C 30 másodperc
primer kötődés 55◦ C 30 másodperc
DNS szintézis 72◦ C 60 másodperc
extenzió 4 ◦C 4 perc
Az utolsó amplifikációs körben alkalmazott reakcióelegy és primerek a 8. és 9.
táblázatban vannak összefoglalva. A reakció paraméterek az előző ampifikációs kör
paramétereivel egyeztek meg. A reakciót Perkin Elmer Gene Amp 2400 PCR System
készülékkel (Perkin Elmer Cetus, Foster City, CA, USA) végeztük. Pozitív és negatív
kontrollokat egyaránt használtunk. A termékeket 2%-os agaróz gélen futtattuk (11.
ábra).

54
8. táblázat. A második amplifikációs körben használt reakcióelegy összetevői.
Komponensek
Térfogat
A komponenst gyártó cég
10x PCR puffer 5 µl Roche J0051
Taq Gold enzim 0,5 µl Roche G0448
10 mM dNTP mix 1 µl Roche D 8220
HCV-3 primer 1 µl Genset 351332
HCV-4 primer 1 µl Genset 351334
dH2O 38,5 µl Eppendorf 0032006159
RNS templát 3 µl
Összesen 50 µl
9.táblázat. A HCV detektálása során használt primerek Primer Orientáció Szekvencia (5’-3) GI lokalizáció amplikon
HCV 8 forward ccatagatcactcccctgtg 13-32
HCV 11 reverse gcacggtctacgagacct
2764397 321-304
308
HCV 3 forward ctgtgaggaactactgtcttcacgcag 28-54
HCV 4 reverse cggttccgcagaccactatg
7669491 141-122
114
11. ábra. HCV vírus detektálása humán máj mintákban. Futtatás 2%-os agaróz
gélen. +k– pozitív kontroll, -k – negatív kontroll, 903, 951- tumoros májszövet; 904,
952- tumormentes májszövet. L - DNS létra.
100 bp

55
4.5.10. RNS expresszió detektálása in situ hibridizációval
Az in situ hibridizációs technika adott nukleinsav-szekvenciák kimutatására használt
molekuláris biológiai módszer. Alapja, hogy az ismert egy szálú szekvencia, esetünkben
RNS – egy szakasza és a hozzá tervezett komplementer, jelzett próba adott
körülmények között hibridizálni képesek egymással.
A 644 bp hosszú patkány matrilin-2 fragmentet RT-PCR-el amplifikáltunk a következő
primerekkel (5’ cctaccccaacggcataca 3’), (5’ tgtgtgaacagtggcgaatc 3’). Az amplifikált
fragmentnek pBluescript IISK vektorba történt a klónozása. A sense és antisense
matrilin-2 riboprobákat digoxigenin-UTP-vel jelöltük (Roche, Mannheim, Germany).
6 µm vastag RN-áz mentesen kezelt paraffinba ágyazott metszetek deparaffinálása 3 x
15 percig xilolban, majd 2x10 percig etanolban történt. Ezt követően a metszeteket RN-
áz mentesen készített PBS-ben mostuk. Ezután pepszines emésztés következett (10
µl/ml, 37◦C-on, 25 percig 0,2N HCl-ban). A metszeteket 4%-os paraformaldehydben
(PBS-ben) posztfixáltuk (jégen 10 percig) és 0,25 %-os ecetsavanhidriddel acetiláltuk.
A metszetekre prehibridizáló oldatot tettünk és hibridizálás előtt denaturáltuk a
ribopróbákkal együtt (94◦C- 4 perc). A prehibridizáló oldatot kicseréltük az ugyanilyen
összetételű, de jelzett próbát is tartalmazó hibridizáló elegyre, amit 53◦C-on történő
inkubálás követett 12-16 órán át.
Negatív kontrollként párhuzamos metszeteken az antiszenz RNS próba helyett szenz
próbát hibridizáltunk. A hibridizációt mosások követték: 20 percig a hibridizációs
hőmérsékleten 2x nátrium-citrate/klorid pufferben (SSC); majd 20µg/ml RNA-ázzal
kezeltük TNE pufferben 30 percig (10 mM Tris- HCl, pH 7,05; 0,5 M NaCl, 1mM
EDTA, 37 ◦C –on), majd tovább mostuk 50 ◦C-on 2x SSC 0.01%, Triton X-100 és 0.1x
SSC, 0.01% TritonX-100, 5mM EDTA oldatokban 20-20 percig.
A reakció detektálása anti-digoxygenin- alkalikus-foszfatáz- IgG-Fab-ellenes
fragmenttel történt (Roche). Az endogén alkalikus-foszfatáz enzimaktivítást 1 perces
20%-os ecetsavval történő kezeléssel blokkoltuk. Az alkalikus-foszfatáz reakció
detektálását NTB (nitro-tetrazolium-blue chloride) (Sigma) és BCIP (5-bromo-4-chloro-
3-indolyl-phosphate) (Sigma) elegyével végeztük. A metszeteket Kaiser- féle glicerin-
zselatinnal fedtük le.

56
4.5.11. Fehérje izolálás
A humán és patkány mintákból a Western blothoz fehérjét izoláltunk.
4.5.11.1. Patkány májakból A fehérje kinyeréshez használt izoláló puffer (0. 1 M NaCl, 50 mM Tris-HCl, 10 mM
EDTA (pH 7.4), proteáz inhibitorok: 2 mM phenylmethylsulfonyl fluorid, 2mM N–
ethylmaleimid, 0.1 M ε-amino-kapronsav, 15 mM benzamidin és 1660 egység
GORDOX (aprotinin)) 1 ml-éhez 100 mg szövetmintát adtunk és Turrax
homogenizátorral végeztük el a szövetek homogenizálást. A homogenizálás után a
mintákat 15 percig 4◦C-on 8000 rpm-en centrifugáltuk, majd a pelletet ugyanilyen
összetételű homogenizáló oldatban szuszpendáltuk és 4M-os ureával történt kezelés
után a mintákat 30 percig jégen extraháltuk. Az így kapott elegyet ismét 8000 rpm-en
15 perig centrifugáltuk 4◦C-on. A kapott felülúszót használtuk a fehérjék további
vizsgálatához. Felhasználás előtt meghatároztuk az íly módon kapott fehérje oldatok
koncentrációját.
4.5.11.2. Humán májakból A humán minták esetében használt izoláló puffer összetétele eltért a patkány minták
esetében használtaktól (a patkány májmintáknál az intenzívebb proteáz gátlás miatt
többféle proteázgátlót is használtunk). Ebben az esetben az izoláló puffer összetétele a
következő volt: 50 mM/l Tris-HCl (pH 7,6), 100 mM NaCl, 10 mM EDTA, 2 mM N-
ethylmaleimid és 2mM phenylmethylsulfonyl fluorid.
A kapott homogenizátumot 8000 rpm-en 15 percig centrifugáltuk, majd a felülúszót
összegyűjtöttük és a fehérjekoncentráció mérése után SDS-poliakrilamid
gélelektroforézisnek vetettük alá.
A fehérje koncentrácót minden esetben a Qubit Fluorimeterrel (Molecular Probes)
mértük a Quanti-iTM Protein Assay Kit (Molecular Probes, Q33212) leírásának
megfelelően.

57
4.5.12. SDS-poliakrilamid gélelektroforézis
Az izolált fehérjék szeparálása SDS-poliakrilamid (SDS-PAGE) gélelektroforézissel
történt nem redukáló körülmények között. Minden mintából 40 µg fehérjét futtattunk
azonos térfogatban. A minták kihígítása és gélre történő felvitele mintapufferben (0,5
M/l Tris-HCl, (pH 6.8), 10% (v/v) glycerol, (0,5 v/v) bromophenol kék) történt. A
patkány mintákat 4-8 %-os grádiens gélen, a humán mintákat pedig 4-12%-os grádiens
gélen futtattuk előrefestett molekulasúly markerekkel párhuzamosan (Amersham
Pharmacia Biotech, Little Chalfont, United Kingdom és PageRulerTM Prestained
Protein Ledder, Fermentas). A futtatást először 80 V-on 10 percig, majd 60 V
feszültségen kb. 1 órán át végeztük, BIORAD PowerPac készülékkel. Futtató pufferként
10 x-es Tris/Glicin/SDS puffert (BIORAD, 161-0732) használtunk tízszeresére hígítva.
Minden esetben ugyanabból a mintasorozatból párhuzamosan két-két gélt futtattunk.
Elektroforézis után az egyik gélt Commassi-kék (BIORAD) fehérje markerrel festettük
meg 30 percig. Majd a fehérjék láthatóvá tétele végett a megfestett gélt differenciáltuk
5% metanolt és 7,5% ecetsavat tartalmazó oldattal. Ez abból a célból történt, hogy
ellenőrizzük a fehérjék futását és szeparálódását a gélben. A másik gélt blottoláshoz
használtuk.
4.5.13. Western blot
A futtatás után nitrocellulóz membránra (Protran, Schleicher and Schuell, BIORAD,
CA) blottoltuk a fehérjét, amely 4°C-on, 10 percig 60 V-on, majd 50 percig 80 V -on
történt. A blottolás után a membránt desztillált vízzel mostuk, majd Ponceau festéssel
ellenőriztük, hogy a fehérjék transzferje megtörtént-e. A festék membránból történő
eltávolításához NaOH-t használtunk, majd mostuk TBS-sel. A filtert ezután egész
éjszakán keresztül blokkoltuk a nem specifikus kötődés megakadályozása miatt. A
blokkoló oldat 5%-os zsírmentes tejpor volt TBS-ben oldva (pH = 7,6). A filtert ezután
nyúl anti-matrilin-2 antitesttel (1: 500 hígítva a blokkoló pufferben) 1 éjszakán keresztül
inkubáltuk 4◦C-on. TBS-Tweennel történő alapos mosás után a membránt ezen antitest
ellen termelt, peroxidáz-konjugált kecske anti-nyúl IgG ellenanyaggal 1: 1000 hígítva

58
ugyancsak a blokkoló oldatban inkubáltuk 1 órán át, végül a jelet erősített
kemilumineszcenciával (ECL, Amersham Pharmacia Biotech) mutattuk ki.

59
5. EREDMÉNYEK
5.1. Matrilin-2 fehérje expresszió vizsgálata patkány májban
immunhisztokémiával
5.1.1. Matrilin-2 vizsgálata immunhisztokémiával normál patkány májban
Normál, kezeletlen patkány májban immunhisztokémiai reakcióval a matrilin-2 a
portális térben, az erek és az epeutak BM-ban, valamint az azt körülvevő kötőszöveti
rétegben volt kimutatható (12. ábra). Az ereket erős pozitivitás jellemezte, míg az
epeutak BM-ja nem mutatott intenzív reakciót, helyenként az epeúti pozitivitás igen
gyenge vagy fragmentált volt. A májparenchymában a szinuszoidok mentén nem volt
matrilin-2 expresszió. A centrális vénákban sem detektáltunk matrilin-2-t.
12. ábra. A, B. Matrilin-2 immunreakció normál patkány májban. Pozitív reakció
detektálható a portális tér (PT) ereiben (nyíl), az epeutakat körülvevő
kötőszövetben (nyíl). Nagyítás: A. 200x; B. 600x.
A
PT
B
PT

60
5.1.2. Matrilin-2 immunhisztokémiai kimutatása májregenerációban
A matrilin-2 expressziójában bekövetkező változásokat először a parciális
hepatectomiát (PH) követően vizsgáltuk F-344 patkány májban. Mint arról már az
irodalmi áttekintésben szó esett, ilyenkor a májtömeg pótlása döntően a hepatociták
osztódása réven történik. A hepatociták DNS szintézisének csúcsa patkányban jól
reprodukálhatóan 24 órával a műtét után van, amit a nem parenchymális sejtek
osztódása 24-48 óra múlva követ. Ennek alapján a parciális hepatectomiával kezelt
patkányokból immunhisztokémiai vizsgálatokhoz a mintákat a PH-t követően a 24., 48.
és a 72.-ik órában leölt állatokból nyertük. Mindhárom időpontban az immunreakció
hasonló pozitivitást mutatott mind a lokalizáció mind pedig az intenzitás tekintetében.
Immunhisztokémiával a különböző időpontokban vett mintákból nem volt
fénymikroszkóppal látható eltérés. A PH utáni metszeteken végzett immunreakciót
összehasonlítva a kezeletlen normál májakból nyert mintákkal a reakció jelentős
hasonlóságot mutatott: a matrilin-2 hasonló lokalizációját figyelhettük meg mint a
normál májakban, azaz minden esetben detektálható volt a matrilin-2 jelenléte a portális
térben (13. ábra). Az epeutak és az erek BM-jában is láttunk hasonlóan gyengén pozitív
reakciót, itt az epeutak BM-jában detektálható pozitív reakció sokkal egyértelműbb volt
mint a kezeletlen májakban. Míg a máj szinuszoidjaiban és a centrális vénákban
ezekben az esetekben sem volt immunhisztokémi módszerrel kimutatható matrilin-2
fehérjeexpresszió.

61
13. ábra. Matrilin-2 fehérje immunhisztokémiai detektálása parciális
hepatectomiával kezelt patkány májakon, PT-portális tér. A nyíl a pozitív reakciót
jelzi. A. 24 órával a PH után, B. 48 órával a PH után, C, D. 72 órával a PH után.
Nagyítás: A, B, C 200x; D 600x.
A következő lépésben a matrilin-2 fehérje expresszióját immunhisztokémiával az ovális
sejtek részvételével történő májregenerációban vizsgáltuk. Az ovális sejtek
proliferációját AAF/PH kezeléssel értük el az „Anyag és módszer” fejezetben leírtaknak
megfelelően. Az AAF/PH kísérleti rendszerben történő májregenerációt a 14. ábra
mutatja be.
A
PT
PT
C D
PT
B
PT

62
14. Ábra. Az AAF/PH kísérleti rendszerben lezajló májregenerációt bemutató
sematikus ábra. Alacsony dózisú modell: az ovális sejt duktuszok a májlebenybe
penetrálnak és szimultán „kis hepatocitákká” differenciálódnak. Magas-dózisú modell:
Az ovális sejtek mélyen a májparenchymába hatolnak és csak jelentéktelen mértékben
változnak. A differenciálódó sejtek fókuszokat képezve tekervényes és elágazó
duktuszokba rendezőnek. A napok száma a parciális hepatectomia után eltelt időt jelenti
(Paku és mtsai 2004).
Kísérleteinkhez a magas – dózisú modellt használtuk. Az ovális sejtek a PH utáni 5.
napon jelentek meg először, számuk ezután a napok múlásával progresszíven
növekedett. A mintavétel az AAF/PH kezelés során a PH-t követően a 9., 11., és 13.
napokon történt. A különböző időpontokban történt mintavételt abból a célból végeztük,
hogy meghatározzuk azt az időpontot, amikor az ovalis sejt proliferáció a
legintenzívebb. Az AAF/PH kezelést követően az ovális sejtek proliferációja és a
duktuláris reakció legintenzívebben a parciális hepatectomiát követően a 13. napon volt
detektálható (15., 16. ábra).
Magas dózisú AAF
Alacsony dózisú AAF
3-4 nap 5-6 nap
8-10 nap 11-13 nap
Norm máj

63
15. ábra. HE festés AAF/PH kezelést követő regenerálódó patkány máj metszeten.
A. Ovális sejt proliferációban gazdag portális tér (nyilak), 200 x; B. Ovális sejtek
(nyilak), 600x
Az ovális sejtek detektálása specifikus OV-6 antitesttel is megtörtént fagyasztott
metszeteken. OV-6-al egyértelműen azonosíthatóak az ovális sejt proliferációban
gazdag portális terek és a májparenchymába mélyen behatoló “ovális sejt képezte
duktuszok”, az ún. „ovális sejt lécek” (16. ábra).
16. ábra. Hepatikus ovális sejtek detektálása OV-6 antitesttel (zöld),
immunfluoreszcenciával AAF/PH kezelt patkány májakon. A. Erős pozitív reakció
a portális terekben (nyíl), 200x; B. Ovális sejt lécek (*), 400x. A kék festődés a
magokat jelzi.
B
A B
A
*
*
B

64
A matrilin-2 fehérje kifejeződése ez esetben erősen követte az OV-6 festődést. A
normál és PH- kezelt májakhoz hasonlóan az AAF/PH regenerációs modellben is
pozitív reakció jellemezte a portális tereket, amelynek intenzitása sokkal erősebb volt,
mint a hepatociták osztódásával történő májregeneráció során és a normál májban.
Továbbá matrilin-2 reakció volt megfigyelhető a szinuszoidok egy részében (a lebenyek
külső 2/3-ban) és az ovális sejt képződményekben, a duktuszokban. Míg a
májparenchyma centrális zónájában - amelyik területen ovális sejtek nem jelentek meg
– a matrilin-2 expressziója nem volt kimutatható (17. ábra).
17. ábra. A matrilin-2 immunhisztokémiai detektálása AAF/PH kezelt patkány
májakon. A. Erős pozitivitás látható a periportális területen, a proliferáló ovális
sejtek területén, a portális térben és a szinuszoidok külső 2/3-ában, 100x. B. Nincs
pozitív reakció a centrális vénában (CV) és a centrolobuláris régió
szinuszoidjaiban, 200x. C. Erős matrilin-2 expresszió detektálása a portális térben
(PT), 400x, AEC. D. Negatív kontroll, 100x. A nyilak a pozitív reakciót jelzik.
C
PT
CV
B
D
A

65
Abból a célból, hogy megvizsgáljuk a matrilin-2 expresszója hogyan követi a laminin
expresszióját, párhuzamos metszeteken minden esetben laminin immunreakciót is
végeztünk. A laminin expressziójának változását már előző kísérletekben leírták (Paku
és mtsai 2004). Ezen eredmények alapján feltételezhettük, hogy a matrilin-2
expressziója hasonló mintázatot mutat, mint a lamininé. Megfigyelhető volt a laminin és
a matrilin-2 koexpressziója (18. ábra).
18. Ábra. Laminin immunhisztokémiai detektálása AAF/PH kezelt patkány
májakon, paraffinba ágyazott metszeten. A. Erős pozitivitás látható a periportális
(nyilak) területen, a proliferáló ovális sejtek területén, a portális térben és a
lebenyke külső 2/3-ának a szinuszoidjaiban, 100x. B. Nincs pozitív reakció a
centrális vénában (CV, nyíl) és centrális szinuszoidokban, 200x, AEC.
D
B CV
A

66
5.2. Matrilin-2 fehérje expresszió detektálása AAF/PH kezelést követően Western
blot technikával
Azonos mennyiségű normál patkány, PH kezelt és AAF/PH kezelt patkány májszövet
homogenizátumot 4-8%-os poliakrilamid gélen futtatva igen jelentős eltéréseket
találtunk az AAF/PH kezelt májakban a normál májak matrilin-2 fehérje
expressziójához képest. Jelentős mennyiségű matrilin-2 volt detektálható az AAF/PH
kezelt májakban, míg a normál májakban illetve a PH-kezelt patkány májakban a
fehérje igen jelentéktelen mennyiségben volt jelen, vagy egyáltalán nem volt
kimutatható.
19. ábra. Matrilin-2 fehérje expresszió detektálása Western blot technikával. PH
kezelt máj (p), AAF/PH kezelt máj (a), kontroll máj homogenizátum (c), uterus,
pozitív kontroll (u); A. A gél Commassie-kékkel festett fehérjeképe. B. A
párhuzamos minták immunoblotja. A matrilin-2 monomer ~ 100kDa-nál
detektálható.
A máj homogenizátum Commassie-val visszafestett fehérjeképén jól látható, hogy a
különböző mintákból azonos mennyiségű fehérjét vittünk fel (19.A.ábra). Több
próbálkozás után a patkány májmintákból a detektált jel mindegyik mintában
megközelítőleg 100 kDa-nál volt látható, ez jól megegyezik a matrilin-2 monomer
A B

67
molsúlyával. Pozitív kontrollként a matrilin-2 fehérjében igen gazdag uterusból nyert
fehérje kivonatot használtuk. Ezzel többek közt az is volt a célunk, hogy ellenőrizzük az
általunk használt antitest specificitását, valamint azt, hogy a szérum felismeri-e a
redukálatlan uterus mintákban már régebben kimutatott oligomereket is (Piecha és mtsai
1999). A matrilin-2 sok más fehérjéhez hasonlóan, amelyekben van von Willebrand
Faktor modul, tartalmaz egy úgynevezett MIDAS motívumot (Metal Ion Dependent
Adhesion Site, fémionfüggő adhéziós hely), amely - nevéből adódóan - fémion kötésre
képes, és ezen keresztül más fehérjékhez kapcsolódhat. Ez a kölcsönhatás
megszüntethető volt azzal, hogy EDTA-t adtunk az extrakciós pufferbe. Az ureát
tartalmazó oldattal pedig megszüntettük az ionos kölcsönhatások nagy részét, illetve az
úgynevezett gyenge kölcsönhatásokat.
Az ábrán jól látható, hogy az uterus szövetkivonatból futtatott mintában a fő csík
elektroforetikus mobilitása a patkány mintákban található monomerrel azonosan fut. Az
uterus mintában látható, hogy a fehérjének több formája is létezik (a proteolízis
megakadályozására proteázgátlót, fenil-metil-szulfonil-fluoridot raktunk a pufferekbe,
ezért a csíkok nem lehetnek a vizsgálat során esetlegesen képződő fehérje-degradáció
melléktermékei). Az uterus kivonatban megjelenő többféle matrilin-2 oligomer
molsúlya megfelel a más szerzők által már azonosított méreteknek (Piecha és mtsai
2002; Korpos és mtsai 2005). Azonban a fehérjének ezeket a formáit a patkány májban
többszöri próbálkozás után és intenzív proteáz gátlás mellett sem sikerült azonosítani.
Azt hogy az oligomerek hiánya egy intenzív proteolitikus emésztés eredménye a
májban, vagy más folyamatok játszanak szerepet ebben a jelenségben, nem tudjuk.
A kísérletet AAF/PH kezelt májakon több egymást követő napon vett mintákból is
megismételtük (9., 11., 13., napokon) Pozitív kontrollként ez esetben patkány vesét
használtunk (20. ábra).

68
20. ábra. Matrilin-2 fehérje expresszió detektálása Western blot technikával. 1, 2, 3
normál májak; 4, 5, 6 - AAF/PH kezelt minták a PH-t követő 9, 11, és 13. napon
vett mintákból, 7- pozitív kontroll, vese. A matrilin-2 monomer csík ~ 100kDa-nál
detektálható. NK-negatív kontroll, az elsődleges antitest helyett blokkoló pufferrel
inkubáltuk.
A Western blottal az eltérő napokon vett mintákból nem volt kimutatható eltérés a
matrilin-2 fehérje mennyiségében. A Western blottal nyert eredmények alátámasztották
az immunhisztokémiával nyert eredményeket: a matrilin-2 fehérje nagy mennyiségben
detektálható az ovális sejtek proliferációjával történő májregeneráció során, míg a
normál májban és a hepatociták részvételével történő májregenerációban nem, vagy
csak alig mutatható ki.
5. 3. Matrilin-2-t termelő sejtek detektálása májregenerációban
A fehérjeexpresszió vizsgálatára irányuló kísérleteink azt sugallták, hogy az AAF/PH
májregenerációs modellben az ovális sejtek proliferációval történő regeneráció során
sokkal kifejezettebb a matrilin-2 fehérje expresszió, mint a hepatociták regenerációja

69
során a PH modellben illetve a normál kezeletlen májakban. Ennek alapján az volt a
célunk, hogy megvizsgáljuk milyen sejtek termelhetik a megemelkedett matrilin-2
fehérjét a májregeneráció során, és megvizsgáljuk, hogy van-e kimutathtaó eltérés
mRNS expressziós szinten a matrilin-2 termelésében a normál májban, és az AAF/PH
kezelt májakban. Mivel a PH-kezelt májak a normál májhoz hasonlóan viselkedtek a
Western blot analízis során, így ezzel a csoporttal mRNS szinten nem foglalkoztunk.
A matrilin-2 fehérjét termelő sejtek azonosítását nem radioaktív in situ hibridizációval
végeztük és a reakciót alkalikus foszfatáz-rendszerrel detektáltuk. Az in situ
hibridizációs kísérletek alapján az intenzíven pozitív jelnek értékelhető indigókék
reakciót csak az ovális sejtek citoplazmájában láttunk, míg az érett hepatociták nem
mutattak pozitivitást (21. ábra). Ennek alapján úgy tűnik, hogy az ovális sejtek termelik
és szekretálják a matrilin-2-t az őket burkoló BM-ba, amely a proliferáció során
képződő „ovális-sejt léceket” veszi körül. Feltételezhető, hogy a matrilin-2-nek fontos
szerepe lehet az ovális sejtek indukálta májregenerációban. Az in situ hibridizáció során
kapott eredmény az immunhisztokémia és Western blot eredményeinket is
alátámasztja.
21. ábra. A, B. Matrilin-2 mRNS detektálása ovális sejtekben in situ
hibridizációval. A periportális területen jól láthatók az indigókék reakciót mutató
ovális sejt duktuszok (nyíl), PT-portális traktus. A 200x; B 400x.
*
B
PT
A
PT

70
5.4. Matrilin-2 expresszió vizsgálata real-time RT-PCR analízissel
Annak igazolására, hogy a matrilin-2-t valóban az ovális sejtek termelik a SE II.
Belgyógyászati Klinika Kutatólaboratóriumában (dr Molnár Béla vezetésével) ovális
sejteket és hepatocitákat disszekáltunk lézer mikrodisszekcióval. Az ovális sejteket az
AAF/PH kezelést követően a 13. napon vett májmintákból disszekáltuk. Az ún. „régi”
hepatocitákat normál kezeletlen patkány májból izoláltuk, illetve az „új” hepatocitákat a
PH műtétet követő regeneráció során képződött érett hepatocitákból nyertük. 3-3
párhuzamos mintával végeztük el a kísérletet. A real-time RT-PCR-el kapott adatok
alapján egyik hepatocita frakcióban sem volt detektálható a matrilin-2 expressziója
mRNS szinten (a matrilin-2 mRNS a 40. ciklusnál sem volt detektálható). mRNS
expressziót csak az ovális sejtekben detektáltunk. A mikrodisszekció az ovális sejtekben
gazdagon proliferáló periportális területről történt. A sejtek eredetét AFP (ovális -sejt
marker) expresszió detektálásával igazoltuk. A PCR reakció eredménye is az in situ
hibridizációval nyert eredményeket támasztja alá, miszerint detektáható matrilin-2
mRNS expresszió csak az ovális sejtekben volt.
22. ábra. Az RT-PCR reakció termékek futtatása 2%-os agaróz gélen. Matrilin-2
(Matn2) mRNS detektálható az ovális sejtekben (1), nincs matrilin-2 PCR termék
a „régi hepatocitákban”(2) és az „új hepatocitákban”. (3). A GAPDH (1, 2, 3) belső
kontrollként szerepelt. L – 100 bp DNS létra.
100 bp

71
5. 5. Matrilin-2 expresszió vizsgálata humán májban
5.5.1. Matrilin-2 fehérje expresszió vizsgálata ép és cirrhotikus humán
májszövetben immunhisztokémiai módszerrel
35 sebészileg rezekált tumormintából és azok megfelelő környező szövetéből nyert,
formalinban fixált, paraffinba ágyazott metszeteken végeztük el az „Anyagok és
módszerek” fejezetben leírtaknak megfelelően az immunhisztokémiai reakciókat.
Minden általunk vizsgálatra kiválasztott normál máj HE metszetén a máj szerkezetére
jellemző normál méretű portális tér, fibrosismentes szerkezet volt látható.
A normál májban (23. ábra) illetve a tumor melletti nem cirrhotikus májszövetben a
matrilin-2 csak a portális térben, az erek és az epeutak BM-jában mutatott erősen pozitív
reakciót. Nem volt azonban kimutatható matrilin-2 fehérje expresszió sem a centrális
vénák körül, sem pedig a szinuszoidokban és hepatocitákban sem. Ezek az eredmények
megegyeznek a patkány normál májmintákban kapott immunhisztokémiai
eredményekkel.
23. ábra. A, B. Matrilin-2 immunhisztokémiai reakció normál májban. A matrilin-
2 festődés jól látható a portális erekben (nyíl) és az epeutakat körülvéve (nyílhegy).
A szinuszoidokban nem látható pozitív reakció. A 200x, B 600x, AEC.
A HCC-k, melyekből a mintavétel történt, 42, 8 %-a cirrhotikus talajon alakult ki. A
tumor környéki cirrhotikus májszövetben a matrilin-2 expressszió fokozott mértékben
BA
PT
PT

72
volt jelen a cirrhotikus nodulusok széli részén, a szinuszoidok mentén. Különösen erős
pozitív reakciót észleltünk a cirrhosisra jellemző kötőszöveti sövényekben, az itt
előforduló erekben és a proliferáló epeutak BM zónájában (24. ábra).
24. ábra. A. Negatív matrilin-2 immunfestés a tumor melletti nem cirrhotikus
szövet acinusaiban, pozitivitás csak a tumor-máj határ kötőszövetében
detektálható, elsősorban az erekben (nyíl) 400x. B. Intenzív matrilin-2 pozitív
reakció a tumor melletti cirrhotikus szövetben, az erekben és a májszinuszoidok
mentén (nyilak), 200x. C. Intenzív matrilin-2 reakció a cirrhotikus kötőszöveti
sövényekben (nyíl), 100x; D. Matrilin-2 reakció cirrhotikus szinuszoidokban
(nyilak) 600x, AEC.
5.5.2. Matrilin-2 fehérje expressziójának vizsgálata hepatocelluláris carcinomában
Az általunk vizsgált, morfológiailag jól differenciált tumorok (G1, grade1) trabekuláris
növekedési mintázatot mutattak. A tumorsejtekre jellemző volt a pleiomorfizmus eltérő,
bár viszonylag enyhe foka. A kevésbé differencált (G2, grade 2) tumorokban a
D
B
C
A

73
daganatsejtekre az acináris, tubulo-acináris szerkezet volt jellemző. Mindegyik
szövetminta tartalmazott ép vagy cirrhotikus peritumorális májszövetet
összehasonlításként. A 35 vizsgált esetből 15 eset cirrhotikus talajon kialakult HCC
volt. A további 20 esetben a tumort környező szövet ép volt vagy enyhe fibrosis
jellemezte. Nodulusok képződése, duktuláris reakció nem volt megfigyelhető ezekben
az esetekben.
Az immunhisztokémiával detektált matrilin-2 megjelenési mintázata a HCC-ben
jelentősen eltért a normál májszövetéhez képest. Igen intenzív matrilin-2 fehérje
expresszió volt megfigyelhető az újonnan képződő az erek BM zónájában, a tumor
neovaszkulatúra mentén. A tumorok differenciáltsági fok szerint nem mutattak eltérést.
A makrotrabekuláris egységekben azonban nem jelent meg matrilin-2 a tumor sejteket
kísérve, hasonlóan a normális máj trabekuláihoz.
*
25. ábra. Matrilin-2 lokalizációja jól és gyengén differenciált tumorokban (nyíl). A.
Heterogén tumor, (*) jól differenciált (G1), (**) mérsékelten (G2) differenciált
területek a HCC-ben. A jól differenciált tumorsejtek trabekuláit matrilin-2
expresszió kíséri (nyíl), a rosszabbul differenciált területeken a nagyobb tumorsejt
csoportokon belül, az egyes sejtek környezetében nincs matrilin-2 expresszió, 200x.
B. Rosszul differenciált tumorban matrilin-2 expresszió csak a
neovaszkulatúrának megfelelően detektálható (G3), 200x.
A matrilin-2 neovaszkuláris BM-ban történő lokalizációjának a megerősítése végett
elemeztük a CD34, mint endothel markert ezeken a tumor mintákon. A CD34
lokalizációban nagy mértékű hasonlóságot mutatott a matrilin-2 expressziójával a HCC-
ben (26. ábra)
* **
A B

74
26. ábra. Immunhisztokémiai reakció trabekuláris (G1) HCC-ben. (A) Matrilin-2,
AEC. (B) CD34 (DAB), 200x. A nyilak a pozitív reakciót jelzik.
5.5.3. A laminin és matrilin-2 kolokalizációjának vizsgálata humán májszövetben
Az irodalmi leírásoknak megfelelően a matrilin-2 különböző struktúrák BM-jában
lokalizálódik és több helyen is kolokalizációt mutatott a lamininnal, például bőrben
(Piecha és mtsai 1999, 2002 a, b). Ezért fontosnak tartottuk, hogy megvizsgáljuk,
hogyan viszonyul a matrilin-2 és a laminin lokalizációja egymáshoz normál májban,
cirrhotikus májban, valamint hepatocelluláris carcinomában.
A 27. ábra a laminin immunhisztokémiai reakcióját mutatja be cirrhotikus májban. A
párhuzamos metszeteken jól látható, hogy a matrilin-2 és a laminin egyes részeken
kolokalizálódik. A részleges kolokalizációt paraffinos metszeten kettős
immunfluoreszcenciával is igazoltuk.
A B

75
27. ábra. Immunhisztokémiai reakció paraffinba ágyazott metszeten. Matrilin-2
(A) és laminin (B) expresszió cirrhotikus nodulusban, 200 x, AEC. C. Laminin és
matrilin kettős immunfloureszcencia (C, D). Zöld-matrilin-2, piros-laminin; sárga-
kolokalizáció, C 200x, D 400x.
5.5.4. Matrilin-2 immunhisztokémiai reakciók morfometriai elemzése és
statisztikai analízise
Az immunhisztokémiai reakciók morfometriai értékelése során a matrilin-2 pozitív
területek eloszlására százalékos arányban a következő eredményeket kaptuk: 0,57 % a
normál májakra, 0,59 % a környező nem cirrhotikus májakra, 3,97 % a cirrhotikus
környező májakra, 4,26 % a cirrhotikus talajon képződött HCC-re és 3,03 % a nem
cirrhotikus talajon képződött HCC-re vonatkoztatva. Ezek alapján azt mondhatjuk, hogy
szignifikánsan több matrilin-2 fehérje volt detektálható a HCC-ben és a cirrhotikus
májakban, mint a nem cirrhotikus környező májakban és az ép szövetekben ( p< 0.0001)
(28. ábra). A cirrhotikus talajon kialakult HCC-k és a nem cirrhotikus talajon képződött
D C
A B

76
HCC-k matrilin-2 fehérje expressziója között nem volt szignifikáns eltérés. Továbbá, a
jól – és közepesen differenciált tumorok sem mutattak szignifikáns eltérést a fehérje
expressziójában a rosszul differenciált tumorokhoz képest. A tumorok
differenciáltságának megfelelően az immunpozitív területek százalékos eloszlására a
következő értékeket kaptuk 3, 78 % G1, 4,62% G2 , 4,33 % a G3 HCC-re ( 29. ábra).
28. ábra. A matrilin-2 immunhisztokémiai reakciók során mért intenzitások
összehasonlítása normál májban, cirrhotikus májban és HCC-ben. NORM =
normál máj; NCIRRH = környező nem-tumoros, nem cirrhotikus májszövet;
CIRRH = környező nem tumoros, cirrhotikus máj; HCC (ncirrh) = nem
cirrhotikus talajon kifejlődött HCC; HCC (cirrh) = cirrhotikus talajon létrejött
HCC. A * a csoportok közötti szignifikáns eltérést jelöli. Az immunhisztokémiai
reakciók analízise a Leica Qwin Pro programmal történt, az adatokat a GrapPad
Prism 2.01 programmal értékeltük ki.
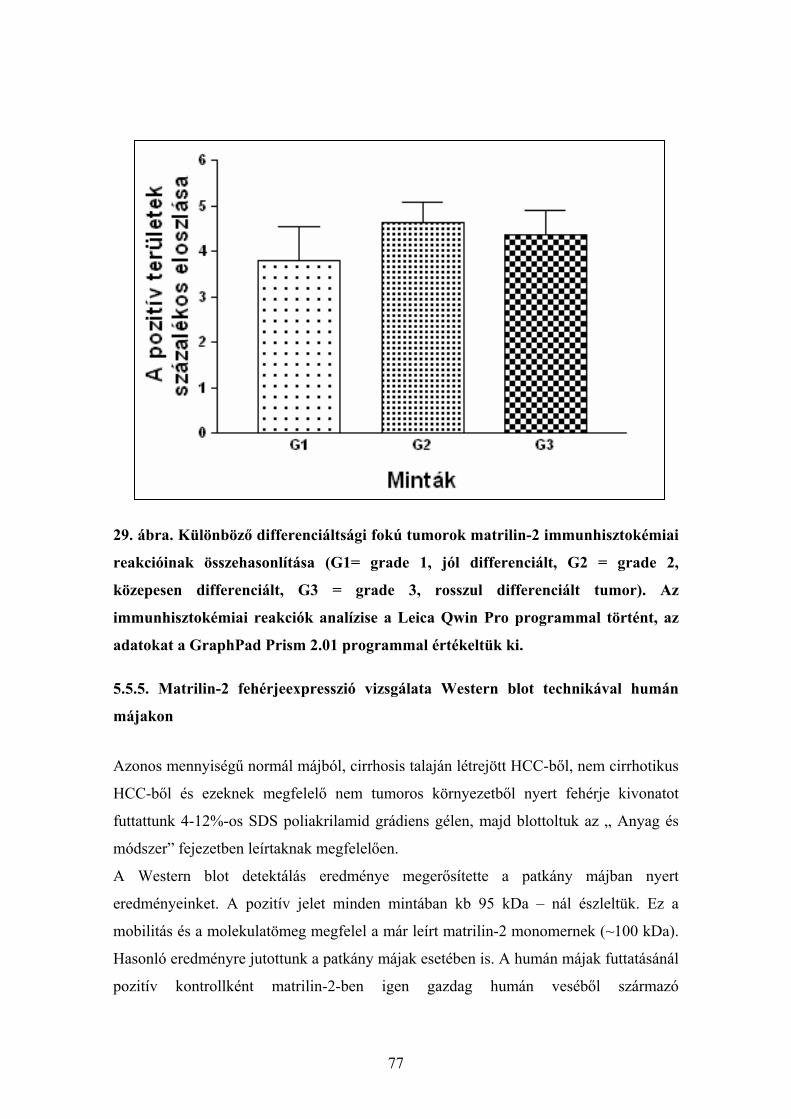
77
29. ábra. Különböző differenciáltsági fokú tumorok matrilin-2 immunhisztokémiai
reakcióinak összehasonlítása (G1= grade 1, jól differenciált, G2 = grade 2,
közepesen differenciált, G3 = grade 3, rosszul differenciált tumor). Az
immunhisztokémiai reakciók analízise a Leica Qwin Pro programmal történt, az
adatokat a GraphPad Prism 2.01 programmal értékeltük ki.
5.5.5. Matrilin-2 fehérjeexpresszió vizsgálata Western blot technikával humán
májakon
Azonos mennyiségű normál májból, cirrhosis talaján létrejött HCC-ből, nem cirrhotikus
HCC-ből és ezeknek megfelelő nem tumoros környezetből nyert fehérje kivonatot
futtattunk 4-12%-os SDS poliakrilamid grádiens gélen, majd blottoltuk az „ Anyag és
módszer” fejezetben leírtaknak megfelelően.
A Western blot detektálás eredménye megerősítette a patkány májban nyert
eredményeinket. A pozitív jelet minden mintában kb 95 kDa – nál észleltük. Ez a
mobilitás és a molekulatömeg megfelel a már leírt matrilin-2 monomernek (~100 kDa).
Hasonló eredményre jutottunk a patkány májak esetében is. A humán májak futtatásánál
pozitív kontrollként matrilin-2-ben igen gazdag humán veséből származó

78
fehérjekivonatot használtunk (30. ábra). Az ábrán jól látható, hogy a matrilin-2 fehérjére
jellemző nagyobb molekulasúlyú oligomer frakció is jelen volt a humán májakban. A
patkány májakban, ezt nem tapasztaltuk, ott csak a matrilin-2 monomert tudtuk
detektálni. Noha ez esetben is ez az oligomer frakció a monomer frakcióhoz képest
elenyésző intenzitású volt és az immunoblot képen alig látható. Megfigyelhető, hogy a
normál humán májban és a tumoros májakban az általunk alkalmazott módszerrel
hasonló intenzitású és azonos mennyiségű matrilin-2 fehérje volt detektálható.
30. ábra. Humán májminták matrilin-2 protein tartalmának összehasonlítása
immunoblot analízis során nyert eredmények alapján. N – normál májból
származó minta; 1 – nem cirrhotikus talajon kialakult HCC-ből származó minta;
2-cirrhotikus talajon létrejött HCC minta; 3 – nem cirrotikus, nem tumoros
környező szövet; 4 – cirrhotikus környező szövet, C- pozitív kontroll, humán vese.
5.5.6. Matrilin-2 mRNS expressziójának real-time RT-PCR analízise humán
májmintákon
A matrilin-2 mRNS expressziót az eddigiekhez hasonlóan humán májmintákban is real-
time RT-PCR módszerrel vizsgáltuk. Olvadáspont analízissel, illetve 2%-os agaróz
gélen történt futtatással ellenőriztük a termékeket.

79
Amint azt már az „Anyagok és módszer” fejezetben írtam, a real-time RT- PCR-rel
kapott adatokat egy relatív kvantifikációra alkalmas program segítségével értékeltük ki.
Ebben a programban az általunk választott GAPDH kontrollgén Ct értékeit a referencia
gén adatainak fenntartott helyre illesztettük be és a kiértékelésnél minden esetben két
csoportot hasonlítottunk össze egymással. A 11. táblázatban a kontroll csoport
elnevezés arra a mintacsoportra vonatkozik, amelyikhez a másik csoportot
hasonlítottuk. (Így például a 11. táblázat 1. sora azt mutatja meg, hogy a cirrhotikus
talajon létrejött HCC-ben a matrilin-2 mRNS expressziója hányszorosára változott a
normál minták esetén tapasztalt értékekhez képest).
A 11. táblázat összefoglalja a real-time RT-PCR eredményeit valamennyi általunk
vizsgált csoportra vonatkoztatva. Ezek az adatok azt mutatják, hogy az egyes
csoportokban jelentősebb, szignifikánsnak mondható mRNS expressziós eltérés a
normál májakhoz képest nem volt detektálható (31. ábra).
Összehasonlítottuk a HCV vírussal fertőzött környező cirrhotikus májak matrilin-2
expresszióját a nem fertőzött környező cirrhotikus májak matrilin-2 expressziójával. A
két csoport között csak jelentéktelen, szignifikánsnak nem mondható eltérést
detektáltunk. A HCV-vel nem fertőzött cirrhotikus májakban a matrilin-2 expressziója
alacsonyabb volt a HCV-vel fertőzött májakhoz képest.
11. táblázat. A matrilin-2 mRNS relatív expressziója real-time RT-PCR-el
detektált Ct értékek alapján. MINTA
CSOPORT KONTROLL CSOPORT
MATRILIN-2 RELATÍV EXPRESSZIÓ
NORM CIRRH(HCC) U 1,21 NS p= 0,756
NORM NCIRRH(HCC) U 1,39 NS p= 0,245
NORM HCC(cirrh) U 1,91 NS p= 0,213
NORM HCC(ncirrh) U 1,55 NS p= 0,245
HCC(cirrh) HCC(ncirrh) D 1,34 NS p= 0,715
CIRRH HCC(cirrh) D 2,32 NS p= 0,415
NCIRRH HCC(ncirrh) U 1,21 NS p= 0,654
HCV(+)cirrh HCV(-)cirrh D 1,14 NS p= 0,785
Magyarázat: A félkövérrel szedett oszlopban feltüntetett csoportok a vizsgálni
kívánt minta csoportot jelentik, amelyet a kontrollként vett csoportokhoz

80
hasonlítottunk (a táblázat 1. oszlopa). U-up-regulált, D-down-regulált relatív
expressziót jelent, NS - nem szignifikáns; NORM - normál máj, HCC -
hepatocelluláris carcinoma, CIRR (HCC) - cirrhotikus talajon létrejött HCC
tumormentes környezete, NCIRRH (HCC) – nem cirrhotikus talajon kifejlődött
HCC tumormentes környezete; HCC (cirrh)-cirrhotikus talajon létrejött HCC;
HCC (ncirrh)- nem cirrhotikus talajon kifejlődött HCC.
31. ábra. A matrilin-2 mRNS relatív expresszióját ábrázoló diagram. A különböző
színnel jelölt csoportok megegyeznek a 11. táblázatban feltüntetett csoportokkal. A
csoportok matrilin-2 mRNS expressziója között nincs szignifikáns eltérés.
-2,5-2
-1,5-1
-0,50
0,51
1,52
1
Matriln-2 mRNS expresszió
NORM CIRRH(HCC)
NORM NCIRRH(HCC)
NORM HCC(cirrh)
NORM HCC(ncirrh)
HCC( cirrh) HCC(ncirrh)
CIRRH HCC(cirrh)
NCIRRH HCC(ncirrh)
HCV(+)cirrh HCV(-)cirrh
MATRILIN-2 mRNS expresszió

81
6. MEGBESZÉLÉS
Az elmúlt évtizedben a normális és kóros szöveti működés sejten belüli eseményeinek
vizsgálata mellett egyre inkább hangsúlyt kaptak azok a kutatások, amelyek a szöveti
kötelék szerves részét képező ECM-re és sejtfelszíni molekulákra irányulnak. Számos
kutatás tárgya a stroma – daganatos sejt kapcsolat, valamint azok mennyiségi, minőségi
változásai összefüggéseinek a vizsgálata.
Munkámban egy nemrégiben felfedezett ECM fehérje, a matrilin-2 expresszióját
tanulmányoztam májszövetben, valamint próbáltam választ keresni arra, hogy mint az
ECM alkotóelemének és a BM komponensének, ennek a fehérjének lehet-e szerepe
kísérletileg indukált májregenerációban, illetve cirrhosisban és HCC-ben.
A tumorok heterogének lehetnek mind a vaszkularizáció mértékében, mind pedig a
stroma jellegében és bizonyos anyagok termelésében egyaránt, többek közt a mátrix
termelésében is. Mivel a daganatok ECM-je különböző módon befolyásolhatja egy
tumor biológiai viselkedését, egyre több olyan irodalmi adat jelenik meg, melyek szerint
bizonyos mátrix komponensek fokozott vagy kóros expressziója rosszabb prognózist
jelenthet (Xiong és mtsai 2006). Szinte minden mátrix fehérje képes a tumoros sejtek
biológiai viselkedésének a befolyásolására.
Matrilin-2 expresszió regenerálódó májban
Jelen tanulmány során a matrilin-2 expresszióját vizsgáltuk normál májakban és
májregenerációs modelleken. Kísérleteinkben ezen újonnan felfedezett mátrix fehérje
csak nagyon kis mennyiségben volt kimutatható a normál patkány májban és a parciális
hepatectomiát követő májregeneráció során. Immunhisztokémiai vizsgálatokkal
kizárólag a portális tér ereiben illetve az epeutak BM zónájában, valamint az ezt
körülvevő kötőszöveti rétegben volt kimutatható matrilin-2 expresszió.
Az ovális sejt indukálta májregenerációban az ovális sejteket magába ágyazó ECM BM
zónájában volt kimutathaó jelentősen megemelkedett matrilin-2 expresszió a normál
májakhoz képest. Ezt mind immunhisztokémiai módszerrel mind Western blot
technikával sikerült igazolni.

82
Kísérleteinkben in situ hibridizációval kimutattuk az mRNS expressziót az ovális sejtek
citoplazmájában. Érett hepatocitákban a matrilin-2 mRNS expressziója nem volt
detektálható. Lézer mikrodisszekcióval disszekált ovális sejtekből és hepatocitákból
tisztított RNS-el végzett PCR kísérletek is alátámasztották az in situ hibridizáció
eredményeit. Ezek birtokában feltételezhető, hogy májregenerációban a vizsgált
modellben, az ECM-ben lezajló változások során a matriln-2-t elsődlegesen termelő
sejtek az ovális sejtek lehetnek.
Ahogyan azt már a bevezetőben is írtam, a matrilin fehérjéket számos szövetben
azonosították (Deák és mtsai 1999, Wagener és mtsai 2005). A matrilin-2–t elsősorban
BM-hoz asszociálva találták meg különböző szervekben (Piecha és mtsai 1999, Segat és
mtsai 2000, Mátés és mtsai 2004, Korpos és mtsai 2005). Az ovális sejt indukálta
májregenerációban különös jelentőséggel bír a BM. A differenciált májsejtekre nem
jellemző a BM. Ezzel szemben a differenciálatlan ovális sejtek BM-on ülnek. A
differenciálódás során ez a struktúra eltűnik (Paku és mtsai 2001). Kísérletileg
igazolták, hogy egy anti-BM ellenes antitest az ovális sejtek igen gyors
differenciálódását idézi elő (Paku és mtsai 2004). Az ECM komponensei intenzíven
termelődnek a regenerálódó májban, ugyanis az ECM normál struktúrája
nélkülözhetetlen a hepatociták differenciált funkciójához. Úgy tűnik, elsősorban a BM
komponenseket tartalmazó mátrix és bizonyos glikoproteinek azok, melyek jelenléte
szükséges ezekhez a folyamatokhoz. Így a laminin és a vele kölcsönható molekulák
jöhetnek először számításba, közöttük lehet a matrilin-2 is (Gressner 1988). A laminin,
ahogy azt már igazolták, kulcsfontosságúnak bizonyult a differenciálódó ovális sejt-
lécek mentén az újonnan formálódó BM-ban (Paku és mtsai 2004). Kísérleteinkben a
matrilin-2 mintázat hasonló lokalizációt mutatott a májregeneráció során a
lamininéhoz, ennek alapján feltételezhető, hogy a matrilin-2 a lamininhez hasonlóan
igen fontos szerepet tölthet be az ECM alakulásában a regeneráció során.
Az ovális sejtek differenciálódását gátló és indukáló mechanizmusok az ECM
komponensei közötti kölcsönhatásukat illetően még nem teljesen ismertek. Az arra
vonatkozó adatok, hogy a regeneráció során egészen pontosan mikor, melyik fázisban
jelennek meg az egyes ECM komponensek még hiányosak. Nemrégiben igazolták egy
kötőszöveti növekedési faktorról, (CTGF = connective tissue growth factor), hogy

83
stimulálja az ECM termelését és kíséletesen indukált májregenerációban számos májsejt
expresszálja, többek közt a máj csillag sejtek és epitheliális sejtek is (Sedlaczek és mtsai
2001). AAF/PH indukált májregenerációs modellben pedig az ovális sejtekben fokozott
expresszióját mérték ennek a növekedési faktornak; továbbá a CTGF up-reguláció
gátlása az ovális sejtek számának szignifikáns csökkenéséhez vezetett. Ez a jelenség is a
kötőszövet igen fontos szerepét támasztja alá az ovális sejtekkel történő
májregenerációban (Pi és mtsai 2005). Feltehető, hogy többek közt ennek a faktornak is
szerepe lehet a matrilin-2 expresszióját fokozó szignalizációs kaszkádban.
Kísérleteink alapján normál patkány májban nem volt kimutatható matrilin-2 expresszió
a centrális vénákban, a szinuszoidokban, és nagyon gyenge expressziót mutatott az
epeutak BM-jában, míg az ovális sejtekkel történő regeneráció során a matrilin-2
expressziója igen intenzív, a szinuszoidok mentén a portális areában, míg a
centrolobuláris régió felé az expresszió eltűnik és a centrális vénákban nem
detektálható a matrilin-2. Ez a matrilin-2-nek a májregeneráció korai szakaszában
betöltött szerepét sejteti.
Az eddig leírtaknak megfelelően (lásd Bevezetés) a máj csillagsejtek lehetnek az ECM-
et felépítő molekulák fő forrásai (Martinez -Hernandez és mtsai 1995, Bedossa és
Paradis 2003). Noha egyre több sejt kap szerepet a máj ECM komponenseinek a
termelésében, még mai napig sem tisztázott teljesen, hogy egyes mátrix komponenseket
milyen sejtek termelnek (lásd Bevezető). Ahogyan már említettem, eredményeink
alapján a matrilin-2–t a regenerálódó májban az ovális sejtek csoportja termelheti. Az a
kérdés, hogy milyen más sejtek, így elsősorban a csillagsejtek, fibroblastok milyen
körülmények között, milyen arányban termelhetnek matrilin-2-t (pl. cirrhosisban)
további vizsgálatok tárgya.
Igazolták, hogy matrilin-2 mRNS expresszió kimutatható hámsejtekben, de a fehérje
expresszió csak szubepitheliális szövetekben volt kimutatható. Patkány bőrben a
dermisz felső rétegében, az erekben és az idegekben volt detektálható (Piecha és mtsai
1999). Humán bőrben in situ hibridizációval keratinocytákban és fibroblastokban is
sikerült matrilin-2 mRNS-t detektálni. A matrilin-2 fehérje termelésének mechanizmusa
és transzportja a BM-on kersztül a stromába pontosan nem ismert, de feltehetőleg a
matrilin-2-vel specifikusan kölcsönható molekulák biztosítják a fehérje transzportját a

84
kötőszöveti stroma irányába és a BM-ban lévő specifikus kötő ligandjai eredményezik a
depozicióját a BM-ba (Piecha és mtsai 2002 a).
A vWFA-szerű doménjének köszönhetően a matrilin-2 a kollegénekkel,
proteoglikánokkal, a laminin család tagjaival és egyéb ECM komponensekkel is
kölcsönhatásba lép, de makromolekuláris kölcsönhatások révén szerepe van az ECM
molekulák összeszerelésében, a kollagén fibrilláris hálózat képzésében is (Piecha és
mtsai 2002 b, Mátés és mtsai 2002). A matrilin-2 vWFA doménje révén erősen kötődik
a kollagén rostokhoz, esetleg azok integráns része, bőrben kolokalizációt mutat a
kollagénnel és szilárd fázisú vizsgálatokban kölcsönhatásba lép a fibrilláris
kollagénekkel, fibrillinekkel, fibronektinnel, laminin-nidogén komplexszel (Piecha és
mtsai 2002 a, b). Ezen ismeretek birtokában feltételezhető, hogy a matrilin-2 a normális
májban a portális terület mátrixot alkotó fibrilláris hálózat kialakításában vehet részt,
ahol a kollagén makromolekuláris szerveződését segítheti. A regenerálódó májban ettől
némileg eltérő szerepe lehet. Az eredményeink azt sugallják, hogy a matrilin-2 a
regenerálódó májban az elsőként termelődő ECM komponensek között lehet.
Feltételezhető, hogy a matrilin-2-re, mint az elsők között megjelenő BM komponensre a
laminin, a kollagének, és más molekulák együttes szerveződésében lehet szükség, amely
a májregenerációban a proliferáló ovális sejtek differenciálatlan állapotának a
fenntartásában alapvető szerepet játszhat. A matrilin-2 pontos funkciójának kérdése
azonban a májregeneráció során még további tisztázásra vár.
A szarvas agancsban, mely egy különlegesen csontosodó szerv, modellje a
csontregenerációnak, emlősökben az egyetlen eset, amikor egy teljes szerv
regenerálódik, vizsgálták a matrilin fehérjék expressziójának változását. Ebben a
differenciálódási rendszerben, is a matrilin-2-t igen intenzíven osztódó, még nem
teljesen differenciálódott sejtek markereként azonosították. Emelt protein expressziós
szinten fejeződött ki mesenchyma sejtekben, előporcban és preosteoblastokban. Ezek az
eredmények szolgáltatták az első bizonyítékait annak, hogy a matrilin-2 különböző
szövetek differenciálatlan sejtjeiben is kifejeződik (Segat és mtsai 2000, Korpos és
mtsai 2005). Újabban pedig cornea-limbalis stem sejtekben mutattak ki matrilin-2
expressziót, nidogénnel és agrinnal párhuzamosan (Schlötzer-Schrehardt és mtsai 2007).

85
A matrilin-2 expressziója humán májban
A munkának ebben a részében a matrilin-2 exressziójával foglalkoztunk normál májban,
májcirrhosisban és hepatocelluláris carcinomában.
Vizsgálataink során a matrilin-2 a humán normál máj portális terében, így az epeutak
BM-jában, a portális artériák, és vénák BM-jában volt erős intenzitással kimutatható,
míg a centrális vénában a matrilin-2 immunohisztokémiai módszerrel nem volt
detektálható. A normál máj szinuszoidjaiban sem volt detektálható a matrilin-2
expressszió. A cirrhotikus májban a matrilin-2 fehérje expressziója jelentős mértékben
volt kimutatható a szinuszoidokban. A HCC-ben a matrilin-2 a neovaszkularizáció BM-
jai mentén figyelhető meg. A tumorban a vaszkularizáció során kialakuló erek
biztosítják a tumor tápanyagellátását. A matrilin-2 neovaszkuláris BM lokalizációjának
megerősítésére CD34-ellenes antitesttel végzett immunreakció, mint az endothel
expressziós markere, részben kolokalizációt mutatott a matrilin-2-vel. Noha a matrilin-2
a májrákok neovaszkulaturáját kirajzolva jelenik meg tumorokban, az eltérő
differenciáltságú tumorok matrilin-2 fehérje expressziója között nem volt
szignifikánsnak mondható eltérés. Az utóbbi években derült fény arra, hogy a
daganatos kapillárisok szerkezete eltérő, ennek alapján több BM komponensről
igazolódott, hogy kevésbé differenciált tumorokban eltűnik illetve fragmentálódik,
amely a BM fellazulásával is jár. A szinuszoidális kapillarizáció a tumor
dedifferenciálódása során történik. A tumorsejtek progresszív dedifferenciálódásával,
atipikussá válnak az ECM termelésére, ezáltal a tumorok metasztázis képző hajlamát
növelve (Giannelli és mtsai 2003). A munkacsoportunk által vizsgált agrinról is ez
igazolódott (Kovalszky és matsai 2004, Batmunkh és mtsai 2007). Az hogy a matrilin-2
fehérje expressziója nem mutat jelentősebb mennyiségbeli eltérést az eltérő
differenciáltságú tumorok és a cirrhosis között, azt jelentheti, hogy ez a fehérje már a
cirrhosis során megjelenik és nincs különösebb jelentősége a tumor kialakulásában,
illetve prognózisában. Az is érdekes jelenség, hogy a matrilin-2 expressziójában az
eltérő talajon (cirrhotikus és nem cirrhotikus) kialakult HCC-k között sincs lényeges

86
különbség. Az irodalomból ugyanis ismeretes, hogy a májrákokat befolyásolja, hogy a
tumor cirrhotikus vagy nem cirrhotikus májban alakul ki. Így egyes stroma
komponensek is eltérhetnek az eltérő talajú tumorokban, pl. a syndecan, a decorin és a
már említett agrin expresszióját nem befolyásolja a cirrhosis jelenléte, míg pl. a
glipican-3 expressziójának fokozódása figyelhető meg cirrhosishoz társult tumorokban
(Kovalszky és mtsai 2004). A matrilin-2 expresszióját feltehetőleg nem befolyásolja
nagy mértékben a cirrhosis jelenléte tumorban.
Mindenestre felsorolt eredményeink alátámasztják azt a tényt, miszerint a matrilin-2 az
eddigi irodalmi adatoknak megfelelően elsősorban a BM struktúrákban volt
detektálható (Piecha és mtsai 1999, 2002, Schlötzer-Schrehardt és mtsai 2007),
májrákokban is ezt találtuk.
Számos közlemény született bizonyos BM-t alkotó, ECM komponensek szerepéről a
fibrogenezisben és a különböző tumorok patogenezisében, többek közt a HCC-ben is.
Így például a kollagén IV, a laminin és az integrinek fokozott expresszióját írták le
cirrhotikus májban és HCC-ben (Kin és mtsai 1994). A COMP (oligomer porc mátrix
fehérje) fokozott expresszióját írták le HCC-ben. Erről a fehérjéről ismertették, hogy a
matrilinekkel szerkezeti hasonlóságot mutat és makromolekuláris kölcsönhatásban áll
velük (Xiao és mtsai 2004). A lamininnel és matrilin-2-vel párhuzamos metszeteken
végzett immunhisztokémiai reakciókkal kimutattuk, hogy ezek a fehérjék részben
kolokalizálódnak. Ez is igazolja, hogy a matrilin-2 a lamininhez hasonlóan egy BM
komponens és a lamininhez, valamint más ECM molekulákhoz hasonlóan a
„kapillarizáció” során felszaporodik a cirrhotikus máj szinuszoidjaiban (Kovalszky
2006).
Immunoblottal a matrilin-2 monomert kb 100 kDa-nál detektáltuk, mind a humán, mind
a patkány májak esetében. A nagyobb molekulasúlyú dimer-, és oligomer frakciónak
megfelelő fehérje a patkány májakban nem volt kimutatható, míg minden humán májban
csekély mennyiségben ugyan, de jelen volt ez a frakció is. Eddigi közleményekben
publikált Western blot kísérletek során kapott adatok a matrilin-2-t kimutatták dimer és
oligomer formában is (Piecha és mtsai 2002, Korpos és mtsai 2005). Annak, hogy saját
eredményeink nem ezt mutatták, több oka lehet: a májban olyan intenzív proteolitikus
hasítása történik a fehérjének, hogy csak a monomer formája marad meg; de

87
elképzelhető, hogy a májból csak a matrilin-2 oligomerek mutathatók ki, vagy a
fehérjének ez a formája van túlsúlyban a többihez képest.
Western blottal nem volt kimutatható mennyiségi eltérés a humán normál májban, a
cirrhotikus és nem cirrhotikus környező májakban valamint a HCC-ben. Ez
ellentmondásosnak tűnik az immunhisztokémiával nyert eredményekhez képest, amely
alapján a cirrhotikus májban és a tumorokban is megemelkedik a matrilin-2
mennyisége. A matrilin-2 monomer sávok intenzitásának hasonlósága a különböző
mintákban a cirrhotikus májban található fehérje nem megfelelő szolubilizációjával
magyarázható a normál máj és a nem cirrhotikus májakhoz képest. Egy másik
magyarázat lehetne erre az ellentmondásra, hogy a portális terek képleteinek BM-jai
igen nagy mennyiségű matrilin-2-t tartalmaznak. Az újonnan képződött szinuszoidok
BM-ja viszont relatíve az összmennyiséghez képest csekélyebb mennyiségben tartalmaz
matrilin-2-t. Bár immunhisztokémiával ez egyértelműen detektálható, de ez
mennyiségben elmarad a Western blottal mérhető összehasonlításban.
Összességében eredményeink alapján feltételezhető, hogy a matrilin-2 mind a
májregenerációban mind a kapillarizációban az elsők között megjelenő ECM
komponensként szerepelhet. Elképzelhető, hogy ezekben az esetekben is a már ismert
„adaptor” funkciója révén a mátrix átépítésében vesz részt más fehérjékkel együttes
kölcsönhatásban.
Eredményeink újabb kérdések sorozatát vetik fel:
1) Milyen molekulákkal együtt jelenik meg a matrilin-2 a májregenerációban és a
cirrhosisban, valamint a tumorokban ?
2) Milyen transzkripciós faktorok indukálják az expresszióját?
3) Milyen molekulák hatására csökken a matrilin-2 fehérje expressziója a májban és a
májregeneráció melyik fázisban következik ez be?
A matrilinek carcinogenezisben és fibrosisban betöltött szerepének precíz
tanulmányozása knock-out állatokon lehetséges. Ilyen irányú vizsgálatok már
folyamatban vannak.

88
7. KÖVETKEZTETÉSEK
1. A normál patkány májszövetben a matrilin-2 fehérje a portális terek
ereinek és epeútjainak BM-ban fordul elő, míg a májszinuszoidokban
a matrilin-2 expressziója nem mutatható ki.
2. Az ovális sejtekkel történő májregeneráció során patkányban a
matrilin-2 expresszió megnő a normál májban detektált expresszióhoz
képest és kimutatható a szinuszoidokban valamint az ovális sejt
képezte duktuszokban is. A matrilin-2 mRNS expresszióját az ovális
sejtekben az RT-PCR és az in situ hibridizáció is igazolta, tehát a
matrilin-2 termelésben az ovális sejtek részt vesznek, melyet elsőként
detektáltunk az irodalomban.
3. A matrilin-2 az ovális sejt duktuszok mentén kolokalizációt mutat a
lamininnal, tehát a lamininhoz hasonlóan fontos komponens lehet az
ECM-ben a májregeneráció során.
4. Humán májban elsőként mutattuk ki a matrilin-2 expressziót mind
mRNS, mind fehérje szinten.
5. Normál humán májban fehérje szinten a matrilin-2 az epeutak, a
portális tér ereinek BM zónájában detektálható, a szinuszoidok
mentén nem volt megfigyelhető matrilin-2 fehérje pozitivitás.
6. A cirrhotikus májszövetben a matrilin-2 fehérje expressziót a
proliferáló epeutak és az erek BM zónájában, valamint a
szinuszoidokban azonosítottuk.
7. A hepatocellularis carcinomákban (HCC) erős matrilin-2 expresszió
figyelhető meg az újonnan képződött erek BM-jában, mely azonban
nem mutat szignifikáns eltérést a különböző differenciáltságú
tumorokban.
8. Az endothelt jellemző CD34 fehérje expresszió a HCC-ben részben
kolokalizál a matrilin-2 expresszióval.

89
8. ÖSSZEFOGLALÁS
A hepatocelluláris carcinoma (HCC) az egyik leggyakoribb malignus daganatok közé
sorolható a világon. A hepatociták malignus transzformációja olyan többlépcsős
folyamat eredménye, amelyben a daganat kialakulásához vezető genetikai elváltozások
mellett egyre jobban felismerjük a tumor extracelluláris mátrixának (ECM) jelentőségét,
mely cél-orientált terápia célpontja is lehet.
A mátrix biológia csak az utóbbi évtizedben fedezte fel a nem-kollagén szerkezetű ECM
fehérjékhez tartozó matrilin fehérjék családját. A matrilin fehérjék négy tagja közül a
matrilin-2 szöveti előfordulása a legszélesebb körű: számos szerv bazális membránjában
(BM) leírták jelenlétét. Amikor ebben az értekezésben ismertetett munka elkezdődött,
humán májjal kapcsolatos irodalmi adatok még nem voltak a matrilin-2-ről. Kutatásaink
legfőbb célkitűzése éppen ezért a matrilin-2 expressziójának tanulmányozása volt
májregenerációban, cirrhosisban és HCC-ben, figyelembe véve az ECM jelentőségét
ezen folyamatokban.
Kísérleteinkben tanulmányoztuk a matrilin-2 mRNS és fehérje expresszióját patkány
normál májban, parciális hepatectomiát követően hepatocitákkal történő, illetve ovális
sejtek indukálta májregenerációs kísérleti modelleken. Elsőként mutattuk ki, hogy az
ovális sejtekkel történő májregenerációban az ovális sejtek termelik a matrilin-2-t. Ezt
fehérje és mRNS szinten is igazoltuk, detektáltuk a matrilin-2 monomer formáját.
Humán májmintákon (35 HCC, 15 cirrhotikus, 20 nem cirrhotikus környező szövet) és
10 normál máj) a matrilin-2-t a normál máj portális tereiben, az epeutak és a portális
erek BM zónájában elsőként detektáltuk; a szinuszoidokban nem volt kimutatható
matrilin-2 fehérje expresszió. A cirrhotikus májak szinuszoidjaiban erős matrilin-2
fehérje expressziót mutattunk ki. A HCC-ben intenzív matrilin-2 fehérje expressziót
találtunk az erek mentén, amely a tumorok differenciáltsági fokával nem mutatott
összefüggést. Feltételezhető, hogy a matrilin-2 a májregenerációban az elsők között
megjelenő ECM komponensként szerepelhet. A fokozott matrilin-2 expresszió
cirrhosisban és a tumorokban e fehérjének a kapillarizációban és a
neovaszkularizációban betöltött szerepére utalhat. Elképzelhető, hogy ezekben az
esetekben is a már ismert „adaptor” funkciója révén a daganat mátrix átépítésében vesz
részt, más fehérjékkel együttes kölcsönhatásban.

90
9. SUMMARY
Hepatocellular carcinoma (HCC) is among the most common causes of cancer death
worldwide. The malignant transformatiormation of the hepatocytes is a longterm multistep
process, in which the genetic alterations and molecular changes in the extracellular matrix
(ECM) play role together in carcinogenesis and tumor progression.
The recently described matrilin protein family is part of the ECM, their pathophysiological
role as well as distribution in the liver have not yet been studied. Matrilin-2 displays a broad
tissue distribution and has been detected in basement membranes of other tissues. Considering
its role in cell growth and tissue remodeling we aimed to study the expression of matrilin-2
during liver regeneration, in normal human liver, HCC and surrounding cirrhotic liver.
Liver regeneration was induced in rats by partial hepatectomy (PH) and 2-
acetylaminofluorene (AAF)/partial hepatectomy (PH) in experimental models.
A total of 35 cases of surgically resected HCCs, 35 corresponding surrounding liver tissues
and 10 normal liver samples were used for the study of human tissues. The expression of
matrilin-2 was analyzed both at mRNA and protein levels. Matrilin-2 was detected in normal
rat liver and partially hepatectomized liver in the portal area, but could not be demonstrated in
the acini. In the AAF/PH model the oval cells but not the hepatocytes produced matrilin-2
mRNA. Increase in protein level in the AAF/PH regenerating liver model was demonstrated
by Western blotting. The protein was present in the basement membrane zone around the
tubules formed by oval cells.
In normal human liver matrilin-2 expression was detected in normal liver, portal blood
vessels, while sinusoids were negative. Cirrhotic surrounding tissues of HCCs showed
intensive matrilin-2 staining along the sinusoids. Tumorous neovasculature was found to be
strongly positive in HCCs. No differences, however, were detected by morphometry based on
the grade of HCC.
Taken together, our data show that among mesenchymal cells hepatic oval cells produce
matrilin-2, a novel ECM protein which appears during capillarization in liver cirrhosis and
hepatocarcinogenesis. Based on our findings, it is suggested that matrilin-2 is one of the
important components of ECM during stem cell-driven liver regeneration and is also
synthetised during the remodeling of the cirrhotic ECM and tumor stroma. Matrilin-2 as
putative adaptor protein might help other molecules for proper ECM assembly during the
reorganization of matrix.

91
10. IRODALOMJEGYZÉK
Aizawa Y, ShibamotoY, Takagi I, Zeniya M, Toda G. (2000) Analysis of factors affecting the
appearance of hepatocellular carcinoma in patients chronic hepatitis C. Cancer, 89: 53-59.
Alison M, Golding MH, Sarraf CE. (1996) Pluripotential liver stem cells: facultative stem
cells located in the biliary tree. Cell Prolif, 29: 373-402.
Alison M. (2005) Liver stem cells: implications for hepatocarcinogenesis. Stem Cell Rev, 1:
253-260.
Andrisani OM, Barnabas S. (1999) The transcriptional function of the hepatitis B virus X
protein and its role in hepatocarcinogenesis. Int J Oncol, 15: 373-379.
Argraves WS, Deák F, Sparks K, Kiss I, Goetinck PF. (1987) Structural features of cartilage
matrix protein deduced from cDNA. Proc Natl Acad Sci USA, 84: 464-468.
Arsura M, Cavin LG. Nuclear factor-kappa B and liver carcinogenesis. (2005) Cancer Lett,
229: 157-169.
Aszódi A, Bateman JF, Hirsch E, Baranyi M, Hunziker EB, Hauser N, Bösze Z, Fasller R.
(1999) Normal skeletal development of mice lacking matrilin1: redundant function of
matrilins in cartilage? Mol Cell Biol, 19: 7841-7845.
Aszódi A, Beier DR, Hiripi. L, Bösze Z, Fassler R. (1998) Sequence, structure and
chromosomal localization of Crtm gene encoding mouse cartilage matrix protein and its
exclusion as a candidate for murine achondroplasia. Matrix Biol, 16: 563-673.
Aszódi A, Hauser N, Studer D, Paulsson M, Hiripi L, Bösze Z. (1996) Cloning, sequencing
and expression analysis of mouse cartilage marix protein (CMP) cDNA. Eur J Biochem, 236:
970-977.

92
Aszódi A, Módis L, Páldi A, Rencendroj A, Kiss I, Bösze Z. (1994) The zonal expression of
chicken cartilage matrix protein gene in the developing skeleton of transgenic mice. Matrix
Biol, 14: 181-190.
Aumailley M, Gayraud B. (1998) Structure and biological activity of the extracellular matrix.
J Mol Med, 76: 253-65.
Barba G, Harper F, Harada T, Kohara M, Goulinet S, Matsuura Y, Eder G, Schaff Z,
Chapman MJ, Miyamura T, Brechot C. (1997) Hepatitis C virus core protein shows a
cytoplasmic localization and associates to cellular lipid storage droplets. Proc Natl Acad Sci
USA, 94: 1200-1205.
Batmunkh E, Tátrai P, Szabó E, Lódi C, Holczbauer A, Páska C, Kupcsulik P, Kiss A, Schaff
Z, Kovalszky I. (2007) Comparison of the expression of agrin, a basement membrane heparan
sulfate proteoglycan, in cholangiocarcinoma and hepatocellular carcinoma Hum Pathol, 38:
1508-1515.
Bedossa P, Paradis V. (2003) Liver extracellular matrix in health and disease. J Pathol, 200:
504-515.
Belluoccio D, Schenker T, Baici A, Trueb B. (1998) Characterization of human matrilin-3.
Genomics, 53: 391-394.
Belluoccio D, Trueb B. (1997) Matrilin-3 from chicken cartilage. FEBS Lett, 415: 212-216.
Block TM, Mehta AS, Fimmel CJ, Jordan R. (2003) Molecular viral oncology of
hepatocellular carcinoma. Oncogene, 22: 5093-5107.
Briggs MD. (2004) Missense mutations in the beta strands of the single A-domain of matrilin-
3 result in multiple epiphyseal dysplasia. J Med Genet, 41: 52-9.

93
Buckner JH, Wu JJ, Reife RA, Terato K, Eyre DR. (2000) Autoreactivity against matrilin-1 in
a patient with relapsing polychondritis. Arthritis Rheum, 43: 939-43.
Buendia MA. (1992) Hepatitis B viruses and hepatocellular carcinoma. Adv. Cancer Res, 59:
167-226.
Buendia MA. (2000) Genetics of hepatocellular carcinoma. Semin Cancer Biol, 10: 185-200.
Canen EH, Yamada KM. (2001) Fibronectin, integrins and growth control. J Cell Physiol,
189: 1-13.
Chapman KL, Mortier GR, Chapman K, Loughlin J, Grant ME, Briggs MD. (2001) Mutations
in the region encoding the von Willebrand factor A domain of matrilin-3 are associated with
multiple epiphyseal dysplasia. Nat Genet, 28: 393-396.
Chen Q, Johnson DM, Haudenschild DR, Tondravi MM, Goetinck PF. (1995) Cartilage
matrix protein forms a types II collagen-independent filamentous network: Analysis in
primary cell cultures with a retrovirus expression system. Mol Biol Cell, 6: 1743-1753.
Chen Q, Zhang Y, Johnson DM, Goetinck PF. (1999) Assembly of a novel cartilage matrix
protein filamentous network: molecular basis of differential requirement of von Willebrand
factor A domains. Mol Biol Cell, 10: 2149-2162.
Chiquet-Ehrismann R. (2004) Tenascins. Int J Biochem Cell Biol, 36: 986-990.
Chisari FV. (2000) Viruses, immunity, and cancer: Lessons from hepatitis B. Am J Path, 156:
1118-1132.
Clarke B. (1997) Molecular virology of hepatitis C virus. J Gen. Virol, 78: 2397-2410.
Craig JR. (1997) Cirrhosis, hepatocellular carcinoma, and survival. Hepatology, 26: 798-799.

94
Deák F, Piecha D, Bacharati C, Paulsson M, Kiss I. (1997) Primary structure and expression
of matrilin-2, the closest relative of cartilage matrix protein within the von Willebrand factor
type A module superfamily. J Biol Chem, 272: 9268-9274.
Deák F, Wagener R, Kiss I, Paulsson M. (1999) The matrilins: a novel family of oligomeric
extracellular matrix proteins. Matrix Biol, 18: 55-64.
Deng-Fu Yao, Zhi-Zhen Dong, Min Yao (2007): Specific molecular markers in hepatocellular
carcinoma, Hepatobiliary Pancrat Dis Int, 6: 241-247.
Dumble ML, Croager EJ, Yeoh GC, Quail EA. (2002) Generation and characterization of p53
null transformed hepatic progenitor cells: oval cells give rise to hepatocellular carcinoma.
Carcinogenesis, 23: 435-445.
Desmet VJ. (2005) Cystic diseases of the liver. From embryology to malformations.
Gastroenterol Clin Biol, 29: 858-860.
Eleazar JA, Memeo L, Jhang JS, Mansukhani MM, Chin S, Park SM, Lefkowitch JH, Bhagat
G. (2004) Progenitor cell expansion: an important source of hepatocyte regeneration chronic
hepatitis J Hepatol, 41: 983-991.
Evarts RP, Nagy P, Marsden ER, Thorgeirsson SS. (1987) A precursor-product relationship
exists between oval cells and hepatocytes in rat liver. Carcinogenesis, 8: 1737-1740.
Falkowski O, An HJ, Ianus IA, Chiriboga L, Yee H, West AB, Theise ND. (2003)
Regeneration of hepatocyte 'buds' in cirrhosis from intrabiliary stem cells. J Hepatol, 39: 357-
364.
Farber E. (1956) Similarities in the sequence of early histological changes induced in the liver
of the rat by ethionine, 2-acetylaminofluorene, and 3’ methyl-4-dimethylaminoazobenzene.
Cancer Res, 16: 142-148.
Fausto N. (2001) Liver regeneration: from laboratory to clinic. Liver Transpl, 7: 835-844.

95
Fausto N. (2004) Liver regeneration and repair: hepatocytes, progenitor cells, and stem cells.
Hepatology, 39: 1477-1487.
Fiegel HC, Glüer S, Roth B, Rischewski J, von Schweinitz D, Ure B, Lambrecht W, Kluth D.
(2004) Stem-like cells in human hepatoblastoma. Histochem Cytochem, 52: 1495-501.
Forbes S, Vig P, Poulsom R, Thomas H, Alison M. (2002) Hepatic stem cells. J Pathol, 197:
510-518.
Frank S, Schulthess T, Landwehr R, Lustig A, Mini T, Jenö P, Engel J, Kammerer RA. (2002)
Characterization of the matrilin coiled-coil domains reveals seven novel isoforms. J Biol
Chem, 277: 19071-19079.
Friedman SL. (2004a) Mechanism of disease: mechanisms of hepatic fibrosis and therapeutic
implications. Nature Clin Practice Gastroenterol Hepatol, 1: 98-105.
Friedman SL. (2004b) Stellate cells: a moving target in hepatic fibrogenesis. Hepatology, 40:
1041-1043.
Fujita M, Furukawa H, Hattori M, Todo S, Ishida Y, Nagashima K. (2000) Sequential
observation of liver cell regeneraton after massive hepatic necrosis inauxiliary partial
orthotopic liver transplantation. Mod Pathol, 13: 152-157.
Füle T, Baghy K, Tátrai P, Péterfia B, Kovalszky I. (2006) A stroma szerepe a daganatok
biológiai viselkedésében. Orvosképzés, 3. 137-244.
Füle T, Kovalszky I. (2002) Vírusok molekuláris diagnosztikája. Magy Onkol, 46: 17-22.

96
Gelse K, Poschl E, Aigner T. (2003) Collagens-structure, function, and biosynthesis. Adv
Drug Deliv Rev. 55: 1531-1546.
Gervain J, Simon G Jr, Papp I, Szabóné BK. (2001) A magyarországi krónikus C
vírushepatitises betegek vírustípus- és szubtípus meghatározása. Orv Hetil, 2001. 142: 1315-
1319.
Giannelli G, Antoniaci S. (2000) Biological and clinical revelance of Laminin-5 in cancer.
Clin Exp Metastasis, 18: 439-443.
Giannelli G, Quaranta V, Antonaci S. (2003) Tissue remodelling in liver diseases. Histol
Histopathol, 18: 1267-1274.
Gressner AM. (1998) The cell biology of liver fibrogenesis - an imbalance of proliferation,
growth arrest and apoptosis of myofibroblasts. Cell Tissue Res, 292: 447-52.
Grisham JW. (1980) Cell types in long term propagable cultures of rat liver. Ann NY Acad
Sci, 349: 128-134.
Hauser N, Paulsson M, Heinegard D, Morgelin M. (1996) Interaction of cartilage matrix
protein (CMP) with aggrecan. Increased covalent cross-linking with tissue maturation. J Biol
Chem, 271: 32247-32252.
Hauser N, Paulsson M. (1994) Native cartilage matrix protein (CMP). A compact trimer of
subunits assembled via a coiled-coil α-helix. J Biol Chem, 269: 25747-26753.
Hayashi J. Aoki H, Arakawa Y, Hino O. (1999) Hepatitis C virus and hepatocarcinogenesis.
Intervirology, 42: 205-210.
Higgins GM, Anderson RM. (1931) Experimental pathology of the liver: restoration of the of
the white rat following partial surgical removal. Experimental Pathol, 12: 186–202.
Hodkinson PS, Mackinnon AC, Sethi T. (2007) Extracellular matrix regulation of drug
resistance in small-cell lung cancer. Int J Radiat Biol, 28. 1-9.

97
Hofseth LJ, Saito S, Hussain SP, Espey MG, Miranda KM, Araki Y, Jhappan C, Higashimoto
Y, He P, Linke SP, Quezado MM, Zurer I, Rotter V, Wink DA, Appella E, Harris CC. (2003)
Nitric oxide-induced cellular stress and p53 activation in chronic inflammation. Proc Natl
Acad Sci USA, 100: 143 -148.
Hsia CC. Evarts RP. Nakatsukasa N, Marsden E, Thorgeirsson SS. (1992) Occurence of oval-
type cells in hepatitis B virus associated human hepatocarcinogenesis. Hepatology, 16: 1327-
1333.
Jackson GC, Barker FS, Jakkula E, Czarny-Ratajczak M, Mäkitie O, Cole WG, Wright MJ,
Smithson SF, Suri M, Rogala P, Mortier GR, Baldock C, Wallace A, Elles R, Ala-Kokko L,
MD Briggs. (2004) Missense mutations in the β strands of the single A-domain of matrilin-3
result in multiple epiphyseal dysplasia. J Med Genet, 41: 52-59.
Kabosova A, Azar DT, Bannikov GA, Campbell KP, Durbej M, Ghohestani RF, Jones JC,
Kenney MC, Koch M, Ninomiya Y, Patton BL, Paulsson M, Sado Y, Sage EH, Sasaki T,
Sorokin LM, Steiner-Champliaud MF, Sun TT, Sundarraj N, Timpl R, Virtanen I, Ljubimov
AV. (2007) Compositional Differences between Infant and Adult Human Corneal Basement
Membranes. Invest Ophthalmol Vis Sci, 48: 4989-99.
Kalluri R. (2003) Basement membranes: structure, assembly and role in tumor angiogenesis.
Nat Rev Cancer, 3: 422-433.
Katyal S, Oliver JH 3rd, Peterson MS, Ferris JV, Carr BS, Baron RL. (2000) Extrahepatic
metastases of hepatocellular carcinoma. Radiology, 216: 698-703.
Kim TH, Wendy M, Mars DB, Stolz B, Petersen E, Michalopoulos GK. (1997) Extracellular
matrix remodeling at the early stages of liver regeneration in the rat. Hepatology, 26: 896-904.
Kin M, Torimura T, Ueno T, Inuzuka S, Tanikawa K. (1994) Sinusoidal capillarization in
small hepatocellular carcinoma. Pathol Int, 44: 771-778.

98
Kiss A, Lotz G, Kaposi NP, Schaff Zs. (2002) Hepatitisvírusok és hepatocarcinogenesis. Orv
Hetil, 143: 83-86.
Kiss I, Deák F, Holloway RG, Delius H, Mebust KA, Frimberger E, Argraves WS, Tsonis
PA, Winterbottom N, Goetnick PF. (1989) Structure of the gene for cartilage matrix protein, a
modular protein of the extracellular matrix. J. Biol. Chem, 264: 8126-8134.
Klatt AR, Nitsche DP, Kobbe B, Mörgelin M, Paulsson M, Wagener R. (2000) Molecular
structure and tissue distribution of matrilin-3, a filament-forming extracellular matrix protein
expressed during skeletal development. J Biol Chem, 275: 3999-4006.
Klatt AR, Nitsche DP, Kobbe B, Macht M, Paulsson M, Wagener R. (2001) Molecular
structure, procesing, and tissue distribution of matrilin-4. J Biol Chem, 276: 17267-17275.
Klatt AR, Paulsson M, Wagener R. (2002) Expression of matrilins during maturation of
mouse skeletal tissues. Matrix Biol, 21: 289-296.
Ko YP, Kobbe B, Paulsson M, Wagener R. (2005) Zebrafish (Danio rerio) matrilins: shared
and divergent characteristics with their mammalian counterpart. Biochem J, 386: 367-379.
Korpos E, Molnar A, Papp P, Kiss I, Orosz L, Deak F. (2005) Expression pattern of matrilins
and other extracellular matrix proteins characterize distinct stages of cell differentiation
during antler development. Matrix Biol, 24: 124-135.
Kovalszky I, Baghy K, Tátrai P, Schaff Zs. (2006) A májfibrogenezis pathomechanizmusa.
Orv Hetil, 33: 1593-1599.
Kovalszky I, Dudás J, Gallai M, Hollósi P, Tátrai P, Tátrai E, Schaff Z. (2004)
Proteoglikánok a májban. Magy Onkol, 48: 207-213.
Kovalszky I. (1993) Máj fibrogenesis-fibrózis, cirrhózis. Orv Hetil, 1: 59-64.

99
Lee JS, Thorgeirsson SS. (2006) Comparative and integrative functional genomics of HCC.
Oncogene, 25: 3801- 3809.
Leighton MP, Nundlall S, Starborg T, Meadows RS, Suleman F, Knowles L, Wagener R,
Thornton DJ, Kadler KE, Boot-Handford RP, Briggs MD. (2007) Decreased chondrocyte
proliferation and dysregulated apoptosis in the cartilage growth plate are key features of a
murine model of epiphyseal dysplasia caused by a matn3 mutation. Hum Mol Genet, 16:
1728-1741.
Lengyel G, Tulassay Z. (2007) A májtranszplantáció után visszatérő hepatitis-C-vírus-fertőzés
kezelése. Orv Hetil, 148: 1875-1881.
Li H, Fan X, Houghton J. (2007) Tumor microenvirovment: the role of the tumor stroma in
cancer. J. Cell Biochem, 101: 805-815.
Llovet J, Ricci S, Mazzaferro V, Hilgard P, Raoul J, Zeuzem S, Poulin-Costello M, Moscovici
M. (2007) Randomized phase III trial of sorafenib versus placebo in patients with advanced
hepatocellular carcinoma. ASCO Annual Meeting Proceedings, 25:1.
Lowes KN, Brennan BA, Yeoh GC, Olynyk JK. (1999) Oval cell numbers in human chronic
liver diseases are directly related to disease severity. Am J Pathol, 154: 537-541.
Mabuchi A, Haga N, Maeda K, Nakashima E, Manabe N, Hiraoka H, Kitoh H, Kosaki R,
Nishimura G, Ohashi H, Ikegawa S. (2004) Novel and recurrent mutations clustered in the
von Willebrand factor A domain of MATN3 in multiple epiphyseal dysplasia. Hum Mutat,
24: 439-40.
Maher JJ, McGuire RF. (1990) Extracellular matrix gene expression increases preferentially
in rat lipocytes and sinusoidal endothelial cells during hepatic fibrosis in vivo. J Clin Invest,
86: 1641-1648.

100
Mann HH, Özbek S, Eugel J, Paulsson M, Wagener R. (2004) Interactions between the
cartilage oligomeric matrix protein and matrilins. Implications for matrix assembly and the
pathogenesis of chondrodysplasisas. J Biol Chem, 279: 25294-25298.
Mannhalter C, Koizar D, Mitterbauer G. (2000) Evalution of RNA isolation metods and
reference genes for RT-PCR analyses of rare target RNA. Clin Lab Med, 38: 171-177.
Martinez-Hernandez A, Amenta PS. (1993) The hepatic extracellular matrix. Ontogenesis,
regeneration and cirrhosis. Virchows Archiv A Pathol Anat, 423: 77-84.
Martinez-Hernandez A, Amenta PS. (1995) The extracellular matrix in hepatic regeneration.
FASEB J, 9: 1401-1410.
Martinez-Hernandez A, Mertinez- Delgado F, Amenta PS. (1991) The extracellular matrix in
hepatic regeneration. Localization of collagen types I, III, IV, laminin, and fibronectin. Lab
Invest, 64: 157-166.
Mátés L, Korpos E, Déak F, Liu Z, Beier DR, Aszódi A, Kiss I. (2002) Comparative analysis
of the mouse and human genes (Matn2 and MATN2) for matrilin-2, a filament-forming
protein widely distributed in extracellular matrices. Matrix Biol, 21: 163-174.
Mátés L, Nicolae C, Mörgelin M, Deák F , Kiss I , Aszódi A. (2004) Mice lacking the
extracellular matrix adaptor protein matrilin-2 develop without obvious abnormalities. Matrix
Biol, 23: 195-204.
Merz H, Malisius R, Mannweiler S, Zhou R, Hartmann W, Orscheschek K, Moubayed P,
Feller AC. (1995) ImmunoMax. A maximized immunohistochemical method for the retrieval
and enhancement of hidden antigens. Lab Invest, 73: 149-156.

101
Mundlos S, Zabel B. (1994) Develpomental expression of human cartilage matrix protein.
Dev Dyn, 199: 241-252.
Nagy P, László V, Schaff Zs. (2006) A hepatocarcinogenesis folyamata humán májban. Magy
Onkol, 50: 107-112.
Nagy P. (1995) The facultative stem cell: a new star in liver pathology. Pathol Oncol Res. 1:
123-126.
Nagy P, Teramoto T, Factor VM, Sanchez A, Schnur J, Paku S, Thorgeirsson SS. (2001)
Reconstitution of liver mass via cellular hypertrophy in the rat. Hepatology, 33: 339-345.
Okimura A, Okada Y, Makihira S , Pan H , Yu L , Tanne K, Imai K, Yamada H, Kawamoto
T, Noshiro M, Yan W, Kato Y. (1997) Enhancement of cartilage matrix protein synthesis in
arthritic cartilage. Arthritis Rheum, 40: 1029-1036.
Paku S, Schur J, Nagy P, SS Thorgeirsson. (2001) Origin and strctural evolution of the early
proliferating oval cells in rat liver. Am J Pathol, 158: 1313-1323.
Paku S, Nagy P, Kopper L, Thorgeirsson SS. (2004) 2-acetylaminofluorene dose-dependent
differentiation of rat oval cells into hepatocytes: confocal and electron microscopic studies.
Hepatology, 39: 1353-1361.
Pan OH, Beck K. (1998) The C-terminal domain of matrilin-2 assembles into a three-stranded
α-helical coiled coil. J Biol Chem, 273: 14205- 14209.
Paulsson M, Heinegard D. (1979) Matrix proteins bound to associatively prepared
proteoglycans from bovine cartilage. Biochem J, 183: 539-545.
Paulsson M, Heinegard D. (1981) Purification and structural caracterization of a cartilage
matrix protein. Biochem J, 183: 539-545.

102
Paulsson M, Heinegard D. (1982) Radioimmunoassay of the 148-kilodalton cartilage protein.
Biochem J, 207: 207-213.
Petersen BE, Zajac VF, Michalopoulos GK. (1998) Hepatic oval cell activation in response to
injury following chemically induced periportal or pericentral damage in rats. Hepatology, 27:
1030-1038.
Pfaffl MW, Horgan GW, Dempfle L. (2002) Relative expression software tool (REST) for
group-wise comparison and statistical analysis of relative expression results in real-time PCR.
Nucleic Acids Res, 1: 30-36.
Pi L, Oh SH, Shupe T, Petersen BE. (2005) Role of the connective tissue growth factor in
oval cell response during liver regeneration after 2-AAF/PHx in rats. Gastroenterology, 128:
2077-2088.
Piecha D, Muratoglu S, Morgelin M, Hauser N, Studer D, Kiss I, Paulsson M, Deak F. (1999)
Matrilin-2, a large, oligomeric matrix protein, is expressed by a great variety of cells and
forms fibrillar networks. J Biol Chem, 274: 13353-13361.
Piecha D, Hartmann K, Kobbe B, Haase I, Mauch C, Krieg T, Paulsson M. (2002a)
Expression of matrilin-2 in human skin. J Invest Dermatol, 119: 38-43.
Piecha D, Wiberg C, Morgelin M, Reinhardt DP, Deak F, Maurer P, Paulsson M. (2002b)
Matrilin-2 interacts with itself and with other extracellular matrix proteins. Biochem J, 367:
715-721.
Roskams T, Yang SQ, Koteish A, Durnez A, DeVos R, Huang X, Achten R, Verslype C,
diehl AM. (2003) Oxidative stress and oval cell accumulation in mice and humans with
alcoholic and nonalcoholic fatty liver disease. Am J Pathol, 163: 1301-1311.

103
Roskams T. (2006) Liver stem cells and their implication in hepatocellular and
cholangiocarcinoma. Oncogene, 25: 3818-3822.
Roskams TA, Theise ND, Balabaud C, Bhagat G, Bhathal PS, Bioulac-Sage P, Brunt EM,
Crawford JM, Crosby HA, Desmet V, Finegold MJ, Geller SA, Gouw AS, Hytiroglou P,
Knisely AS, Kojiro M, Lefkowitch JH, Nakanuma Y, Olynyk JK, Park YN, Portmann B,
Saxena R, Scheuer PJ, Strain AJ, Thung SN, Wanless IR, West AB. (2004) Nomenclature of
the finer branches of the biliary tree: canals, ductules, and ductular reactions in human livers.
Hepatology, 39: 1739-1745.
Rougier P, Mitry E, Barbare JC, Taieb J. (2007) Hepatocellular carcinoma (HCC): An
Update. Semin Oncol, 4: 12-20.
Sadlier DM, Connolly SB, Kieran NE, Roxburgh S, Brazil DP, Kairaitis L, Wang Y, Harris
DC, Doran P, Brady HR. (2004) Sequential extracellular matrix-focused and baited-global
cluster analysis of serial transcriptomic profiles identifies candidate modulators of renal
tubulointerstitial fibrosis in murine adriamycin-induced nephropathy. J Biol Chem, 279:
29670 -2980.
Santoni-Rugiu, E, Jelnes, P, Thorgeirsson, SS, Bisgaard, HC. (2005) Progenitor cells in liver
regeneration: molecular responses controlling their activation and expansion. APMIS, 113:
876-902.
Sarraf C, Lalani EN, Golding M, Anilkumar TV, Poulsom R, Alison M. (1994) Cell behavior
in the acetylaminofluorene-treated regenerating rat liver. Light and electron microscopic
observations. Am J Pathol, 145: 1114-1126.
Saxne T, Heinegard D. (1989) Involvment of non-articular cartilage, as demonstrated by
release of a cartilage-specific protein, in rheumatoid arthritis. Arthritis Rheum, 32: 1080-
1086.
Schaff Z, Nagy P. Pathology techniques and grading systems in the diagnosis of HCC. In
Liver Cancer. Eds. Okuda K, Tabor E. Churchill Livingstone, London, 1997: 111-133.

104
Schaff Z, Sárosi I, Hsia CC, Kiss A, Tabor E. (1995) p53 in malignant and beningn liver
lesions. Eur J Cancer, 31A: 1847-1850.
Schaff Zs, Illyés Gy. A máj és az epeutak patológiája. In: Kopper L és Schaff Zs (szerk),
Patológia. Medicina, Budapest, 2004: 827-839.
Schaff Zs, Székely E, Járay B, Kiss A. (2004) A májdaganatok patológiája. Orv Hetil, 145:
368-374.
Schaffener F, Popper H. Capillarization of hepatic sinusoids in man. Gastroenterology, 1963.
44: 239-242.
Schlötzer-Schrehardt U, Dietrich T, Saito K, Sorokin L, Sasaki T, Paulsson M, Kruse FE.
(2007) Characterization of extracellular matrix components in the limbal epithelial stem cell
compartment. Exp Eye Res, nyomtatásban.
Sedlaczek N, Jia JD, Bauer M, Herbst H, Ruehl M, Hahn EG, Schuppan D. (2001)
Proliferating bile duct epithelial cells are a major source of connective tissue growth factor in
rat biliary fibrosis. Am J Pathol, 158: 1239-1244.
Segat D, Frie C, Nitsche PD, Klatt AR, Piecha D, Korpos E, Deák F, Wagener R, Paulsson M,
Smyth N. (2000) Expression of matrilin-1, -2 and -3 developing mouse limbs and heart,
Matrix Biol, 19: 649-655.
Sell S, Salman J. (1984) Ligh- and electron-microscopic autoradiographic analysis of
proliferating cells during the early of chemical hepatocarcinogenesis in the rat induced by
feeding N-2 fluorenylacetamide in a choline-deficient diet. Am J Pathol, 114: 287-300.
Sell S. (2004) Stem cell origin of cancer and differentiation therapy Crit Rev Oncol Hematol,
51: 1-28.

105
Sharma MK , Watson MA, Lyman M, Perry A, Aldape KD, Deak F, Gutmann DH. (2006)
Matrilin-2 expression distinguishes clinically relevant subsets of pilocytic astrocytoma.
Neurology, 66: 127-130.
Staib F, Hussain SP, Hofseth LJ, Wang XW, Harris CC. (2003) TP53 and liver
carcinogenesis. Hum Mutat, 21: 201-16.
Stefánsson SE, Jónsson H, Ingvarsson T, Manolescu I, Jónsson HH, Olafsdóttir G, Pálsdóttir
E, Stefánsdóttir G, Sveinbjörnsdóttir G, Frigge ML, Kong A, Gulcher JR, Stefánsson K.
(2003) Genomewide scan for hand osteoarthritis: a novel mutation in matrilin-3. Am J Hum
Genet, 72: 1448-1459.
Stirpe NS, Goetinck PF. (1989) Gene regulation during cartilage differentiaton: temporal and
spatial expression of link protein and cartilage matrix protein in the developing limb.
Development, 107: 23-33.
Szabo E, Lotz G, Paska C, Kiss A, Schaff Z. (2003) Viral hepatitis: new data on hepatitis C
infection. Pathol Oncol Res, 9: 215-221.
Szabo E, Paska C, Kaposi Novak P, Schaff Z, Kiss A. (2004) Similarities and differences in
hepatitis B and C virus induced hepatocarcinogenesis. Pathol Oncol Res, 10: 5-11.
Taub R. (1996) Liver regeneration in health and disease. Clin Lab Med, 16: 341-360.
Teufel A, Staib F, Kanzler S, Weinmann A, Schulze-Bergkamen H, Galle PR. (2007)
Genetics of hepatocellular carcinoma. World J Gastroenterol, 13: 2271-2282.
Theise ND, Nimmakayalu M, Gardner R, Illei PB, Morgan G, Teperman L, Henegariu O,
Krause DS. (2000) Liver from bone marrow in humans. Hepatology, 32: 11-16.
Thung SM. (1990) The development of proliferating ductular structures in liver disease. Arch
Pathol Lab Med, 114: 407-411.

106
Tsonis PA, Goetinck PF. (1988) Expression of cartilage-matrix genes and localization of their
translation products in the embryonic chick eye. Exp Eye Res, 46: 753-764.
Wagener R, Kobbe B, Paulsson M. (1997) Primary structure of matrilin-3, a new member of a
family of extracellular matrix proteins related to cartilage matrix protein (matrilin-1) and von
Willebrand factor. FEBS Lett, 413: 129-134.
Wagener R, Kobbe B, Paulsson M. (1998 a) Genomic organisation, alternative splicing and
primary structure of human matrilin-4. FEBS Lett, 438: 165-170.
Wagener R, Kobbe B, Paulsson M. (1998 b) Matrilin-4, a new member of the matrilin family
of extracellular matrix proteins. FEBS Lett, 436: 123-127.
Wagener R., Ehlen HW, Ko YP, Kobbe B, Mann HH, Sengle G, Paulsson M. (2005) The
matrilins - adaptor proteins in the extracellular matrix. FEBS Lett, 579: 3323-3329.
Whittaker CA. Hynes RO. (2002) Distribution and evolution of von Willebrand/Integrin A
domains: widely dispersed domains with roles in cell adhesion and elsewhere. Mol Biol Cell,
13: 3369-3387.
Wiberg C, Klatt AR, Wagener R, Paulsson M, Bateman JF, Heinegård D, Mörgelin M. (2003)
Complexes of matrilin-1 and biglycan or decorin connect collagen VI microfibrils to both
collagen II and aggrecan. J Biol Chem, 278: 37698-37704.
Wilson JW, Leduc EH. (1958) Role of cholangioles in restoration of the liver of the mouse
after dietary injury. J Pathol Bacteriol, 76: 441-449.
Winterbottim N, Tondravi MM, Harrington TL, Klier FG, Vertel BM, Goetinck PF. (1992)
Cartilage matrix protein is a component of collagen fibril of cartilage. Dev Dyn, 193: 266-
276.

107
Wu JJ, Eyre DR.(1998) Matrilin-3 forms disulfide-linked oligomers with matrilin-1 in bovine
epiphyseal cartilage. J Biol Chem, 273: 17433-17438.
Xiao Y, Kleeff J, Guo J, Gazdhar A, Liao Q, Cesare PD, Büchler MW, Friess H. (2004)
Cartilage oligomeric matrix protein expression in hepatocellular carcinoma and the cirrhotic
liver. J Gastroenterol and Hepatol, 19: 296-302.
Xiong ZW, Dan C, Guang RX. (2006) Extracelular matrix remodelling in hepatocellular
carcinoma: effects of soil on seed? Medical Hypotheses, 66: 1115-1120.
You LR, Chen CM, Lee YH. Hepatitis C virus core protein enhances NF-kappaB signal
pathway triggering by lymphotoxin-beta receptor ligand and tumor necrosis factor alpha. J
Virol. 1999. 73: 1672-1681.

11. SAJÁT KÖZLEMÉNYEK JEGYZÉKE
Az értekezés alapjául szolgáló elsőszerzős közlemények jegyzéke: Szabó E., Lódi C., Korpos E., Batmunkh E., Rottenberger Z., Deák F., Kiss I, Tőkés AM.,
Lotz G., László V., Kiss A., Schaff Z., Nagy P. Expression of matrilin-2 in oval cells during
rat liver regeneration. Matrix Biol, 2007. 26: 554-560.
IF: 3,679
Szabó E., Korpos É., Batmunkh E., Lotz G., Holczbauer Á., Kovalszky I., Deák F., Kiss I.,
Schaff Zs., Kiss A. Expression of matrilin-2 in liver cirrhosis and hepatocellular carcinoma.
Pathol Oncol Res, 2007. Accepted for publication, august 2007.
IF: 1,241
Szabo E., Lotz G., Paska C., Kiss A., Schaff Z. Viral hepatitis: New data on hepatits C
infection. Pathol Oncol Res, 2003. 9: 215-221.
Szabó E., Páska C., Kaposi Novák P., Schaff Z., Kiss A. Similarities and differences in
hepatitis B and C virus induced hepatocarcinogenesis. Pathol Oncol Res, 2004. 10: 5-11.
Nem az értekezés alapjául szolgáló közlemények:
Paska C., Bögi K., Szilák L., Tőkés A., Szabó E, Sziller I, Rigó J Jr, Sobel G, Szabó I,
Kaposi-Novák P, Kiss A, Schaff Z. Effect of formalin, acetone, and RNA later fixatives on
tissue preservation and different size amplicons by real-time PCR from paraffin-embedded
tissue. Diagn Mol Pathol, 2004. 13: 234-240.
IF 2,292
Lódi C., Szabó E., Holczbauer Á., Batmunkh E., Szíjártó A., Kupcsulik P., Kovalszky I, Paku
S., Illyés G., Kiss A., Schaff Z. Claudin-4 differentiates biliary tract cancers from
hepatocellular carcinomas. Mod Pathol, 2006. 19: 460-469.

109
IF 3,753
Batmunkh E., Tátrai P., Szabó E., Lódi C., Holczbauer A., Páska C., Kupcsulik P., Kiss A.,
Schaff Z., Kovalszky I. Comparison of the expression of agrin, a basement membrane heparan
sulfate proteoglycan, in cholangiocarcinoma and hepatocellular carcinoma. Hum Pathol,
2007. 38: 1508-1515.
IF: 2,81
Borka K., Kaliszky P., Szabó E., Lotz G., Kupcsulik P., Schaff Z., Kiss A. Claudin
expression in pancreatic endocrine tumors as compared with ductal adenocarcinomas.
Virchows Arch, 2007. 450: 549-557.
IF 2,251
Megjelent, illetve elfogadott in extensio közlemények impakt faktora összesen: 16, 026
Az értekezéssel összefüggő, lektorált folyóiratban megjelent idézhető abstractok:
Szabó E., Nagy P., Paku S., Korpos E., Kiss I., Deak F., Lódi C., Páska C., Kiss A., Schaff Z.
High expression of matrilin-2 by stem cells in liver regeneration. 40th Annual Meeting of the
European Association for the Study of the Liver, April 13-17, 2005, Paris, France. J Hepatol,
42. S2, 2005.
Szabó E., Páska C., Deák F., Kiss I., Batmunkh E., Lódi C., Holczbauer Á., Kiss A.,
Kovalszky I., Schaff Z. Matrilin-2, a novel extracellular matrix component in cirrhosis and
hepatocellular carcinoma. 40th Annual Meeting of the European Association for the Study of
the Liver, April 13-17, 2005, Paris, France. J Hepatol, 42. S2, 2005.

110
Szabó E., Páska C., Lódi C., Batmunkh E., Holczbauer Á., Korpos É., Deák F., Kiss I., Kiss
A., Schaff Z. Expression of matrilin-2 in liver cirrhosis and hepatocellular carcinoma. 47th
Annual Meeting of the Hungarian Society of Gastroenterology, Balatonaliga, June 07-11,
2005, Z. Gastroenterol, Issue 5. Volume 43. 2005.
Egyéb kutatási témákból, lektorált folyóiratban megjelent idézhető abstractok:
Szabó E., Kaposi Novák P., Kiss A., Páska Cs., Lotz G., Schaff Zs. Activation of beta-catenin
pathway in human hepatocellular carcinoma negatively correlates to AFP and CD-44
expression. 46th Annual Meeting of the Hungarian Society of Gastroenterology, Balatonaliga,
June 1-5, 2004, Z. Gastroenterol, 42: 438, 2004.
Batmunkh E., Lódi C., Holczbauer Á., Szabó E., Tátrai P., Páska C., Kiss A., Kupcsulik P.,
Kovalszky I, Schaff Z. High expression of agrin in hepatocellular and cholangiocellular
carcinoma. 40th Annual Meeting of the European Association for the Study of the Liver, April
13-17, 2005, Paris, France, J Hepatol, 42. S2, 2005.
Batmunkh E., Lódi C., Holczbauer Á., Szabó E., Tátrai P., Páska C., Kiss A., Kovalszky I.,
Schaff Z. High expression of agrin in hepatocellular and cholangiocellular carcinoma. 47th
Annual Meeting of the Hungarian Society of Gastroenterology, Balatonaliga, June 7-11, 2005.
Gastroenterol, 43, 2005.
Lódi Cs., Szabó E., Batmunkh E., Holczbauer Á., Paku S., Kiss A., Kupcsulik P., Illyés Gy.,
Schaff Zs. High expression of claudin-4 in biliary carcinomas. 20th European Congress of
Pathology, Paris, France, September 3-8, 2005, Virchows Arch, 447: 404, 2005.
Lódi C., Szabó E., Batmunkh E., Holczbauer Á., Kiss A., Illyés G., Paku S., Schaff Z., Nagy
P. High expression of claudin-7 during liver regeneration. 41th Annual Meeting of the
European Association for the Study of the Liver, Viena, Austria, April 26-27, 2006, J
Hepatol, 44. S2, 2006
Lódi C., Szabó E., Batmunkh E., Holczbauer Á., Paku S., Kupcsulik P., Illyés G., Kiss A.,
Schaff Z. Different claudin-4 expression on biliary and hepatocellular carcinomas. 41th

111
Annual Meeting of the European Association for the Study of the Liver, Vienna, Austria,
April 26-27, 2006, J Hepatol, 44. S2, 2006.
Egyéb kutatási témákból készült, nem idézhető abstractok:
Kiss A., Páska C., Szabó E., Holczbauer Á., Szíjártó A., Kupcsulik P., Schaff Z. Markedly
downregulated claudin-4 expression differentiates human hepatocellular carcinomas from
metastatic liver tumors. Falk Symposium, Freiburg, Germany, October 16-17, 2004.
Batmunkh E., Lódi C., Holczbauer Á., Szabó E., Tátrai P, Páska C., Kiss A., Kupcsulik P.,
Kovalszky I., Schaff Z. High expression of agrin in hepatocellular and cholangiocellular
carcinoma. Falk Symposium, Berlin, Germany, October 3-4, 2005.
Lódi C., Szabó E., Holczbauer Á., Batmunkh E., Kiss A., Illyés G., Paku S., Schaff Z., Nagy
P. High expression of claudin-7 during liver regeneration. Falk Symposium, Berlin Germany,
October 3-4, 2005.
Batmunkh E., Tátrai P., Lódi C., Holczbauer Á., Szabó E., Páska C., Kiss A., Kupcsulik P.,
Kovalszky I., Schaff Z. High expression of agrin in cholangiocarcinoma. Falk Symposium,
Freiburg, Germany, October 10-11, 2006.
Lódi C., Szíjártó A., Holczbauer Á., Szabó E., Batmunkh E., Kiss A., Illyés G., Schaff Z.,
Kupcsulik P. High expression of claudin-4 in human cholangiocarcinomas. 6th Congress of
the European Hepato-Pancreato-Biliary Association, Heidelberg, Germany, May 25-28, 2005.
Holczbauer Á., Halász J., Lódi C., Szabó E., Batmunkh E., Benyó G., Galántai I., Kiss A.,
Schaff Zs. Expression of claudins in human hepatoblastoma. 7th Ph.D. Conference at
Semmelweis University, Budapest, Hungary, April 14-15, 2005.
Szabó E., Orbán E., Lódi C., Batmunkh E., Holczbauer Á., Korpos E., Deak F., Kiss I., Kiss
A., Schaff Z. Expression of matrilin-2 in liver cirrhosis and hepatocellular carcinoma. 1st
Central European Regional Meeting, Liver and Pancreas Pathology, Eger, Hungary, june 1-3,
2006.

112
12. KÖSZÖNETNYÍLVÁNÍTÁS
Köszönetet mondok témavezetőmnek, Schaff Zsuzsa professzor asszonynak, aki lehetővé
tette számomra, hogy a vezetése alatt álló Semmelweis Egyetem, II. sz Patológia Intézetben
végezhessem PhD tanulmányaimat. Köszönöm mindenkori támogatását, rendkívüli türelmét,
tanácsait és szakmai segítségét egyaránt. Köszönöm továbbá a részéről jövő kemény, de építő
szavakat is. Szintén szeretném kifejezni köszönetemet dr. Kiss András docensnek, aki
laborvezetőként mindig biztosította számunkra az anyagi és szellemi támogatást egyaránt,
köszönöm a szakmai tanácsait és segítő emberi jó tanácsait. Hálás köszönettel tartozom dr.
Deák Ferencnek és dr. Kiss Ibolyának, akik előkísérletei nélkül ez a munka nem született
volna meg. Külön köszönöm dr. Deák Ferenc szakmai tanácsait, türelmét és hogy lehetővé
tette számomra számos kísérlet elvégzését laboratóriumában. Hálás és baráti köszönettel
tartozom dr. Korpos Évának és Rottenberger Zsoltnak, akik segítettek a kísérletek
elvégzésében. Hasonlóképpen külön köszönöm dr. Nagy Péter egyetemi tanárnak minden jó
tanácsát és szakmai támogatását. Valamint köszönöm dr. Kovalszky Ilona egyetemi
tanárnőnek az értékes szakmai tanácsait. Köszönöm PhD társaimnak, hogy mindig
segítségemre voltak, dr. Lódi Csabának, dr. Szász Attila-Marczellnek, dr. Holczbauer
Ágnesnek, Német Zsuzsannának. Külön köszönettel tartozok dr. Batmunkh Enkjhargal
kollégámnak, akivel végig együtt dolgoztam, és mindig számíthattam rá a segítségnyújtásban.
Köszönöm dr. Tőkés Anna-Máriának a mindenkori bátorító szavait, értékes szakmai
észrevételeit és tanácsait valamint baráti és emberi támogatását. Továbbá köszönettel
tartozom dr. Páska Csilla biológus kollégámnak, aki sokat segített a módszertani alapok
elsajátításában, dr. Lotz Gábor adjunktusnak a türelmét a fotók igényes elkészítésében.
Szeretném kifejezni köszönetemet Szik Ágnesnek, Pekár Zoltánnénak, Azumahné
Erzsébetnek és Oláh Júliának, akik a munkám gyakorlati részében nyújtottak nagy
támogatását. Hasonlóan köszönettel tartozom az intézet minden munkatársának az általuk
nyújtott segítségért, Rigóné Kálé Elvirának az angol nyelvű cikkek lektorálásáért és formába
öntésért tartozom köszönettel.
És végül, de nem utolsósorban szavakba nem önthető hálával tartozom Szüleimnek,
különösképpen Édesanyámnak és Testvéremnek a támogatásukért.





























