Embed Size (px)
Citation preview

Awh. oral Bid. Vol.10, pp.579-584. 1965. Pergamon Press Ltd. Printed in Gt. Britain.
A HISTOCHEMICAL STUDY OF THE SECONDARY CARTILAGE OF THE MANDIBULAR’CONDYLE IN THE
RAT
N. B. B. SYMONS
Dental School, University of St. Andrews, Dundee, Scotland
Summary-The distribution of glycogen, acid mucopolysaccharides, ribonucleic and desoxyribonucleic acids, and alkaline phosphatase activity in the secondary cartilage of the condyle of the mandible in 24-day and l-month-old rats was studied by histo- chemical methods. Comparison was made with the distribution of the same range of substances in primary hyaline cartilage.
There was virtually no difference between primary and secondary cartilage, except that, whereas glycogen was found in all layers of the secondary cartilage in 24day-old rats, in the primary cartilage it was only found in the chondrocytes. In l-month-old rats, however, glycogen distribution was the same as in the primary cartilage. In the primary cartilage mitotic figures were difficult to find even 24 days after birth; in the secondary cartilage, however, they were still numerous in the l-month-old material.
INTRODUCTION
THE TERM secondary or accessory cartilage is employed to describe certain cartilages which are associated with membrane bones and have no topographical connection with, and appear later in development than, the primary cartilaginous skeleton (SCHAFFER, 1930; DE BEER, 1937).
Phylogenetically the primary cartilage of the mammalian chondrocranium is well- established for it can be traced through all the gnathostomes. The status of the secondary cartilages related to the membrane bones of the skull and the clavicle is, however, uncertain in a phylogenetic sense. They are new formations appearing in mammals.
According to Moss (1958), the term “definitive secondary cartilage” should be reserved for those cartilaginous areas which differentiate independently of the primary cartilaginous skeleton and are replaced by endochondral ossification. This cartilage is to be distinguished from those less well organised areas of cartilage which Moss describes as “intermediate”. The latter are not replaced by endochrondral bone forma- tion but are transformed directly into bone.
An additional feature of the definitive secondary cartilages which distinguishes them from “intermediate” secondary cartilage is that they appear at regular and well defined times in foetal development. Examples in man are the condylar, coronoid and symphyseal cartilages of the mandible, the cartilages at the ends of the clavicle, and that for the hamular process of the medial pterygoid plate. It is noteworthy that the largest and longest persisting of these are associated with joints.
579

580 N. B.B. SYMONS
The more transitory areas of secondary cartilage may be simply “an adaptation for resistance to precocious strains and stresses on the part of the rudiments of the membrane bones” (DE BEER, 1937). But it seems unlikely that this explanation is sufficient in the case of, for example, the secondary cartilage of the mandibular condyle which persists until the growth of the jaw is complete.
The most obvious histological difference between primary and definitive secondary cartilage is the relatively scanty amount of intercellular matrix found in the latter, making the secondary cartilage a more cellular tissue.
An additional difference is to be found in their methods of growth. Primary cartilage may grow by apposition or by interstitial growth. Appositional growth can only occur in those areas where there is a perichondrium; it is not found, therefore, in articular or epiphyseal cartilage, except peripherally where this layer is present. In definitive secondary cartilage appositional growth always occurs, for even where it is found in relation to a joint, such as at the condyle of the mandible, a thick layer of fibrous tissue-a perichondrium-forms the actual articular surface.
Since the development and structure of the secondary cartilage of the mandibular condyle had already been studied (SYMONS, 1951, 1952), it was thought that it would be profitable to investigate the histochemical reactions of “definitive” secondary cartilage, as represented by this particular cartilage, and to compare them with the reactions of primary cartilage.
A study of the histochemical reactions of primary cartilage has already been made by PRITCHARD (1952), and his findings have been made use of in this comparison. However, as the primary cartilage examined by PRITCHARD was apparently non- articular, and was entirely foetal in origin, it was decided to supplement his findings by the study of articular cartilage from the post-natal period. The joint selected for this purpose was that between the scapula and the humerus.
MATERIAL AND METHODS
Rats of 2-4 days and I month of age were employed, with a total of fifteen animals from four different litters. The mandibular and shoulder joints were dissected out and fixed in chilled absolute alcohol at 4°C. The material was cleared in benzene and em- bedded in paraffin wax. Undecalcified sections were cut at 10-12~.
Alkaline phosphatase activity was investigated using the simultaneous coupling azo-dye method (PEARSE, 1960). Sodium a-naphthyl phosphate was used as the sub- strate and the diazonium salt of 5 chloro-O-toluidine as the coupling agent; these had previously (SYMONS, 1955) been found to be satisfactory. The incubation times ranged from 10 min to 4 hr; the pH employed was 9.0.
Glycogen was demonstrated by the periodic acid-Schiff method. Sections were flattened out on 70 per cent alcohol with a minimum of heat. After removal of the wax and passing through absolute alcohol they were dipped into a thin (0.25 per cent) solution of celloidin in alcohol-ether. Some sections were incubated with diastase at 37°C as a control, before applying the celloidin coating.
Desoxyribonucleic acid in the nuclei was demonstrated by means of the Feulgen method.

A STUDY OF THE SECONDARY CARTILAGE OF THE MANDIBULAR CONDYLE IN THE RAT 581
Ribonucleic acid distribution was investigated by the use of the pyronin-methyl
green method of TREvAN and SHARROCK (1951) as modified by PEARSE (1960). Control sections were treated with ribonuclease before staining with pyronin-methyl green.
Acid mucopolysaccharides were identified by their affinity for alcian blue at pH 3.0 and methylene blue at pH 3.6.
Some sections were stained with haematoxylin (Mayer’s Haemalum) and eosin in order to observe the general histology for comparison with the sections that had been
subjected to histochemical methods (Fig. 1). The von Kossa silver nitrate method was used in order to demonstrate the distribution of calcium salts.
GENERAL OBSERVATIONS
It has been shown by PRITCHARD (1952) that in relation to the development of
primary cartilage a definite sequence of cell types and matrix can be distinguished. The outermost zone contains perichondrial fibroblasts and tangentially arranged collagen fibres, followed by a zone of small round pre-chondroblasts in a network of
fine fibres, and then by a zone of chondroblasts in which the fibrous matrix is progres- sively masked by a deposit of hyaline cartilage matrix. This zone in turn merges into
definitive cartilage containing chondrocytes and an abundance of hyaline matrix.
An equivalent gradation of cell types can be distinguished in the secondary cartilage of the condyle of the mandible (Fig. 2). The terminology used by PRITCHARD has,
therefore, been employed. It should be noted, however, that this terminology was produced for the description of areas of primary cartilage with a perichondrium and not for articular cartilage.
HISTOCHEMICAL AND CYTOLOGICAL OBSERVATIONS
Alkaline phosphatase
Phosphatase activity was not found in the condylar cartilage until hypertrophic
changes began to occur (Fig. 3). The enzyme activity was then found in the cytoplasm
of the chondrocytes and in the intercellular matrix and quickly reached its maximum. The appearance of the activity in the intercellular matrix was found where the deposi-
tion of inorganic salts was also beginning. The activity remained about the same level in the cartilage until it was replaced by bone formation.
Phosphatase activity in the articular cartilage of primary origin showed an exactly
similar distribution. In primary cartilage which does not ossify, no phosphatase activity was found.
Glycogen
A little glycogen was to be found in the fibrous perichondrium of the condylar cartilage in the 24-day-old rats. A small amount appeared in the prechondroblasts,
and in the chondroblasts and chondrocytes there was a steady increase in the level of storage (Figs 4 and 5). In the latter type of cells the amount of stored glycogen reached the maximum, the cells being packed with granules. In the early stages of hypertrophy of the chondrocytes this intense level of glycogen storage remained but with a few exceptions was lost from the cells as they degenerated.

582 N. B. B. SYMONS
In the l-month-old rats no glycogen was found until the chondrocyte stage was reached (Fig. 6). In the hypertrophic chondrocytes glycogen storage was intense.
The distribution of glycogen in the primary cartilage of the shoulder joint was similar to that of the secondary cartilage in the l-month-old rats.
Feulgen reaction for desoxyribose nucleic acids
The nuclei of the fibroblastic and pre-chondroblastic cells of the condylar cartilage stained deeply with about the same intensity as the mesenchymal cells generally (Fig. 7). In the chondroblasts, however, there was a diminution in the depth of staining which became even more marked in the definitive chondrocytes. This suggests either a loss or a dilution of desoxyribonucleic acids from the series of cells as they reach maturity.
In the articular cartilage of primary origin a similar diminution in the intensity of staining appeared as the cells reached the definitive chondrocyte stage.
Cytoplasmic basophilia and ribonucleic acids
In the condylar cartilage the series of cells from perichondrial fibroblast to chondro- blast showed an increasing cytoplasmic basophilia. With pyronin-methyl green the cytoplasm of the chondroblast stained red whereas the nuclear membrane and chroma- tin stained blue-green. The periphery of the nucleoli stained blue-green and the central area stained red. The basophilia with pyronin shown by the cytoplasm and the nucleoli of the chondroblasts was removed by the action of ribonuclease. This indi- cates that the marked basophilia of these cells is produced by a high content of ribonucleic acid.
In the chondrocytes the level of basophilia decreased sharply and in the region where hypertrophic changes occurred it disappeared completely.
In the articular cartilage from the shoulder joint an increasing cytoplasmic baso- philia was found in passing from the articular surface to the chondroblasts. This level was, however, maintained in the chondrocytes and only dropped off in the zone where marked hypertrophic changes took place.
Mucopolysaccharides
Staining with methylene blue was shown by the matrix of the secondary cartilage of the mandibular condyle; in addition the matrix stained blue-green with alcian blue indicating the presence of acid mucopolysaccharides. With the periodic acid- Schiff test the matrix stained fairly strongly.
Mitotic figures
Mitotic figures were readily found in the secondary cartilage of the condyle both among the perichondrial fibroblasts and the prechondroblasts, but particularly in the latter zone of the cells (Fig. 2). The mitotic figures showed clearly in the Feulgen preparations, but were also distinctly seen in the sections stained with pyronin- methyl green or with haematoxylin and eosin. In both the 2-4day-old rats and the

A STUDY OF THE SECONDARY CARTILAGE OF THE MANDIBULAR CONDYLE IN THE RAT 583
l-month-old animals the mitotic figures were numerous though, as might be expected, they were more numerous in the younger material.
In contrast to this it was difficult to find any mitotic figures in the primary cartilage, even in the 2-4-day-old animals.
DISCUSSION
It is evident that histochemically there is little difference between the secondary cartilage of the condyle of the mandible and primary cartilage, whether the primary cartilage be the post-natal articular variety used in this work or the foetal type studied by PRITCHARD (1952). So far as alkaline phosphatase, desoxyribonucleic acid, ribonucleic acid, and acid mucopolysaccharides are concerned, there is virtually no difference; the distribution of glycogen forms the only exception.
PRITCHARD described alkaline phosphatase activity in the nuclei as well as in the cytoplasm of the enlarging and hypertrophic chondrocytes, whereas in this study it was only found in the cytoplasm. PRITCHARD, however, was employing Gomori’s original calcium-cobalt method whereas the simultaneous azo-coupling dye method had been used in this work. It is now commonly accepted that the phosphatase activity in nuclei, indicated by the older method, is an artifact.
In the very young material from 2-4-day-old rats, glycogen is found in all zones of the secondary cartilage, though only increasing in amount to an intense level as Une moves inward from the articular surface towards the definitive chondrocytes. In the articular cartilage of the shoulder joint no glycogen is found until the zone of chondrocytes is reached, even in the 2-Aday-old rats. In foetal cartilage, according to PRITCHARD, there is no glycogen in either the fibrous perichondrium or in the chondroblasts, though some was found in the prechondroblasts and an intense level in the enlarging chondrocytes.
In the secondary cartilage of the mandibular condyle from the 1 -month-old material, however, glycogen distribution falls into line with that shown by the articular cartilage of primary origin, for it no longer appears until the zone of the definitive chondrocytes is reached.
In the newly born animal there must be considerable activity in the perichondrium of the mandibular condyle for the appositional growth of the secondary cartilage. It seems reasonable to suggest that the glycogen observed in the perichondrium at this stage is related to this growth. The absence of glycogen from this layer in the slightly older animal may be due to the slowing down of appositional growth.
In the light of the work of ELLIOT (1936), the difficulty experienced in finding mitotic figures in the articular cartilage of primary origin is not surprising. ELLICK found that whereas mitotic division occurs in immature cartilage, it is gradually replaced by amitotic division and in the adult animal no mitosis can be seen. Indeed, he stated that in articular cartilage from rats of only l-week-old mitotic figures were very scanty.
In contrast to this the secondary cartilage of the mandibular condyle continued to show mitotic figures throughout the period studied. The secondary cartilage, though in a sense articular, is covered by a thick perichondrium and does not actually form

584 N. B. B. SYMONS
the articular surface. It has been suggested that amitotic division occurs when the need for metabolites exceeds the supply, as is likely to occur in articular cartilage,
where the only nutritional source would appear to be the synovial fluid. In the secon-
dary cartilage, on the other hand, a good nutritional supply can be obtained through the fibrous perichondrium.
It is commonly inferred that the mandibular joint cannot be pressure bearing because of its histological structure; it is possible that absence of pressure has some relation to the continued growth of the condylar cartilage by mitotic division.
R&me-La repartition du glycogene, des mucopolysaccharides acides, des acides ribonucleiques et desoxyribonucleiques et l’activite phosphatasique alcaline ont et&. Ctudiees par des methodes histochimiques dans le cartilage secondaire du condyle mandibulaire de rats, ages de 2 a 4 jours et de I mois. Des comparaisons avec la repartition des memes substances dans le cartilage primaire hyalin ont et6 effect&es.
11 n’y avait pratiquement aucune difference entre les cartilages primaire et secondaire, sauf en ce qui concerne le glycogene. Ce dernier a Cte trouve dans toutes les couches du cartilage secondaire de rats ages de 24 jours, alors que dans le cartilage primaire, ii n’est concentre qu’au niveau des chondrocytes. Dans des rats, ages de 1 mois, la repartition du glycogene est cependant identique a celle du cartilage primaire. Dans ce dernier, des figures de mitose sont rares, meme 2 a 4 jours apres la naissance: dans le cartilage secondaire, elles sont cependant nombreuses chez des rats, ages de 1 mois.
Zusammenfassung-Mit histochemischen Methoden wurde die Verteilung von Glykogen, sauren Mukopolysacchariden, Ribonuklein- und Desoxyribonukleinsaure sowie die Aktivitlt von alkalischen Phosphatasen im sekundaren Knorpel des Unter- kieferkondylus von 2 bis 4 Tage und 1 Monat alten Ratten untersucht. Vergleiche wurden mit der Verteilung der gleichen Substanzen im primaren hyalinen Knorpel angestellt.
Im wesentlichen waren keine Unterschiede zwischen primarem und sekundlrem Knorpel zu finden mit der Ausnahme, dass Glykogen im primaren Knorpel nur in den Chondrozyten gefunden wurde, wahrend diese Substanz in allen Schichten des sekundaren Knorpels bei 2 bis 4 Tage alten Ratten vorhanden war. Bei den 1 Monat alten Ratten war jedoch die Glykogenverteilung die gleiche wie im primaren Knorpel. Hier waren Kernteilungsfiguren selbst 2 bis 4 Tage nach der Geburt schwer zu finden. Tm sekundaren Knorpel waren sie jedoch noch im 1 Monat alten Material zahlreich vorhanden.
REFERENCES
DE BEER, G. R. 1937. The Development of rhe Vertebrate Skull. Oxford University Press. ELLIOTT, H. C. 1936. Studies on articular cartilage. I. Growth mechanisms. Amer. J. Anat. 58, 127-141. Moss, M. L. 1958. Fusion of the frontal suture in the rat. Amer. J. Anut. 102, 141-165. PEARSE, A. G. E. 1960. Histochemistry. (2nd Ed.). Churchill, London. PRITCHARD, J. J. 1952. A cytological and histochemical study of bone and cartilage formation in
the rat. J. Anat. Lond. 86, 259-277. SCHAFFER, J. 1930. Die Stutzgewebe, in: Handbuch der mikroskopischen Anatomic des Menschen
(Ed. by V. MOLLENDORF) 2:2 Springer, Berlin. SYMONS, N. B. B. 1951. Studies on the growth and form of the mandible. Dent. Rec. 71,41-53. SYMONS, N. B. B. 1952. The development of the human mandibular joint. J. Anat., Lond. 86,326332. SYMONS, N. B. B. 1955. Alkaline phosphatase activity in the developing teeth of the rat. J. Anat.,
Lond. 89,238-245. TREVAN, D. J. and SHARROCK A. 1951. A methyl green pyronin-orange G stain for formalin-fixed
tissues. J. Path. Bact. 6.3, 326329.

secti FIG. 1. Sagittal section of the mandibular joint of 2-day-old rat. Undecalcified on. Haematoxylin and eosin. x 40.
tibrc are disc and
FIG. 2. Sagittal section of mandibular condyle. The transition from perichondrial 3blasts, through prechondroblasts, to chondroblasts, is shown. Two mitotic figures present in the prechondroblastic zone, and are indicated by arrows. D, articular ; F, perichondrial fibroblasts; P, prechondroblasts; C, chondroblasts. Haematoxylin eosin. y 330.
PL ATt 1

N. B. B. SYMONS
FIG. 3. Sagittal section of the mandibular joint of 2-day-old rat showing phos- phatase activity. Not counter-stained. The faint colouration of the nuclei is due to a general background staining which is not due to phosphatase activity. Sodium-a- Naphthyl phosphate and 5 chloro-O-toluidine. x 40.
FIG. 4. Sagittal section through the mandibular joint of 2-day-old rat stained by the PAS method. x 40.
PLATE 2
A STUDY OF THE SECONDARY CARTILAGE OF THE MANDIBULAR CONDVLE IN THE RA?
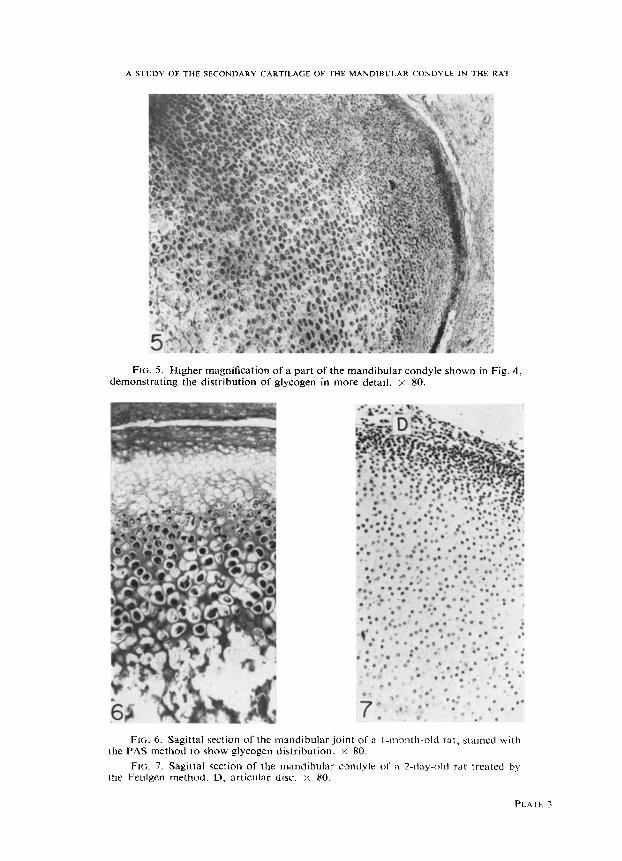
FIG. 5. Higher magnification of a part of the mandibular condyle shown in Fig. 4, demonstrating the distribution of glycogen in more detail. x 80.
FIG. 6. Sagittal section of the mandibular joint of a I-month-old rat, stained with the PAS method to show glycogen distribution. y 80.
FIG. 7. Sagittal section of the mandibular condyle of a 2-day-old rat treated by the Feulgen method. D, articular disc. x 80.
PLATE 3