Embed Size (px)
Citation preview

TECHNOLOGY REPORT
A Fate Map of Tbx1 Expressing Cells Reveals Heterogeneityin the Second Cardiac FieldTuong Huynh,1 Li Chen,1,2 Phillip Terrell,1 and Antonio Baldini1,2,3*1Institute of Biosciences and Technology, Texas A&M University Health Sciences Center, Houston, Texas2Cardiovascular Sciences Program, Baylor College of Medicine, Houston, Texas3Telethon Institute of Genetics and Medicine, Naples, Italy
Received 15 February 2007; Revised 6 May 2007; Accepted 13 May 2007
Summary: Tbx1 is required for the expansion of secondheart field (SHF) cardiac progenitors destined to the out-flow tract of the heart. Loss of Tbx1 causes heartdefects in humans and mice. We report a novel Tbx1Cre
knock-in allele that we use to fate map Tbx1-expressingcells during development in conjunction with a reporterand 3D image reconstruction. Tbx1 descendants consti-tute a mesodermal cell population that surrounds theprimitive pharynx and approaches the arterial pole ofthe heart from lateral and posterior, but not anteriordirections. These cells populate most of the outflowtract with the exception of the anterior portion, thusidentifying a population of the SHF of distinct origin.Both myocardial and underlying endocardial layers werelabeled, suggesting a common origin of these cell types.Finally, we show that Tbx1Cre-positive and Tbx1Cre-neg-ative cell descendants occupy discrete domains in theoutflow tract throughout development. genesis 45:470–475, 2007. Published 2007 Wiley-Liss, Inc.y
Key words: Tbx1; cardiovascular development; secondheart field; Cre-based cell fate mapping; outflow tract of theheart
Tbx1 is critical in many developmental processes (Bal-dini, 2005; Naiche et al., 2005). Its human homolog isdeleted in most cases of DiGeorge syndrome (DGS) andis mutated in some patients with a DGS phenotype(Paylor et al., 2006; Yagi et al., 2003). The phenotypeassociated with Tbx1 loss of function includes, but isnot limited to, developmental abnormalities of headbones and muscles, ear, and cardiovascular system. Thegene is expressed in pharyngeal endoderm and ecto-derm, as well as in the head mesenchyme, pharyngealcore mesoderm, and part of the splanchnic mesoderm(Chapman et al., 1996; Zhang et al., 2005). Expression-based cell fate mapping and conditional deletion demon-strated that Tbx1 is transiently expressed and requiredfor a mesodermal cell population destined to the outflowtract, that is, the second heart field (SHF) (Kelly andPapaioannou, 2007; Xu et al., 2004; Zhang et al., 2006).More complex and diverse functions were identified dur-ing inner ear development, where Tbx1 is not only
required for cell proliferation but also for cell fate deter-mination (Xu et al., 2007). Tbx1-Cre lines have beenimportant tools for providing insights into the develop-mental roles of the gene. Cell fate mapping experimentshave been carried out using three different Tbx1-Crelines by three groups of investigators (Brown et al.,2004; Maeda et al., 2006; Xu et al., 2004). Brown et al.(2004) generated a Tbx1-Cre transgenic line using a7.6 kb sequence (a ligation of four evolutionary con-served segments from a 16.5 kb sequence upstream ofthe Tbx1 coding region). The recombination patternappeared broader than the endogenous expression inthe mesodermal tissues and did not appear to includethe ectodermal domain. Maeda et al. (Maeda et al.,2006) generated a Tbx1-Cre line using a 1.1 kb genomicfragment from the 50 end of the gene. This transgeneincludes only a small portion of the regulatory region ofthe endogenous gene. Indeed, this Cre driver did notinduce recombination in the ectoderm and in the oticepithelium. Xu et al. (2004) described a Tbx1 mutant al-lele, Tbx1mcm, in which a cDNA encoding a Tamoxifen-inducible Cre recombinase had been knocked into theTbx1 locus. While this allele proved useful to addressspecific questions, for example, timed cell fate mapping(Xu et al., 2005), it requires the constant presence ofTamoxifen to reveal transient, highly dynamic expres-sion domains. However, high and prolonged exposure toTamoxifen can cause embryo lethality. In addition, in-ducible Cre recombinases may produce variable resultsby combining the effects of incomplete Cre recombina-tion and incomplete induction.
yThis article is a US Government work and, as such, is in the public do-
main in the United States of America.
* Correspondence to: Antonio Baldini, MD, Institute of Biosciences and
Technology, Texas A&M Health Science Center, 2121 W. Holcombe Blvd.,
Houston, TX 77030.
E-mail: [email protected]
Tuong Huynh and Li Chen contributed equally to this work.
Contract grant sponsor: NIH/NHLBI, Contract grant numbers: HL051524,
HL064832.Published online in
Wiley InterScience (www.interscience.wiley.com).
DOI: 10.1002/dvg.20317
Published 2007 Wiley-Liss, Inc. genesis 45:470–475 (2007)
To maximize the representativeness of expression-driven cell tracing, we generated a new Tbx1
Cre knockin allele. We selected to insert an IRES-Cre construct inthe same site that was used to generate the Tbx1
lacZ
knock-in allele, which recapitulates endogenous geneexpression (Lindsay et al., 2001; Vitelli et al., 2002). Thegene targeting strategy is illustrated in Figure 1a,b.
We crossed Tbx1Cre/þ animals with the R26R reporter
line to reveal Cre activity. We found evidence of recom-bination in presomite embryos harvested at E7.5-E8. At
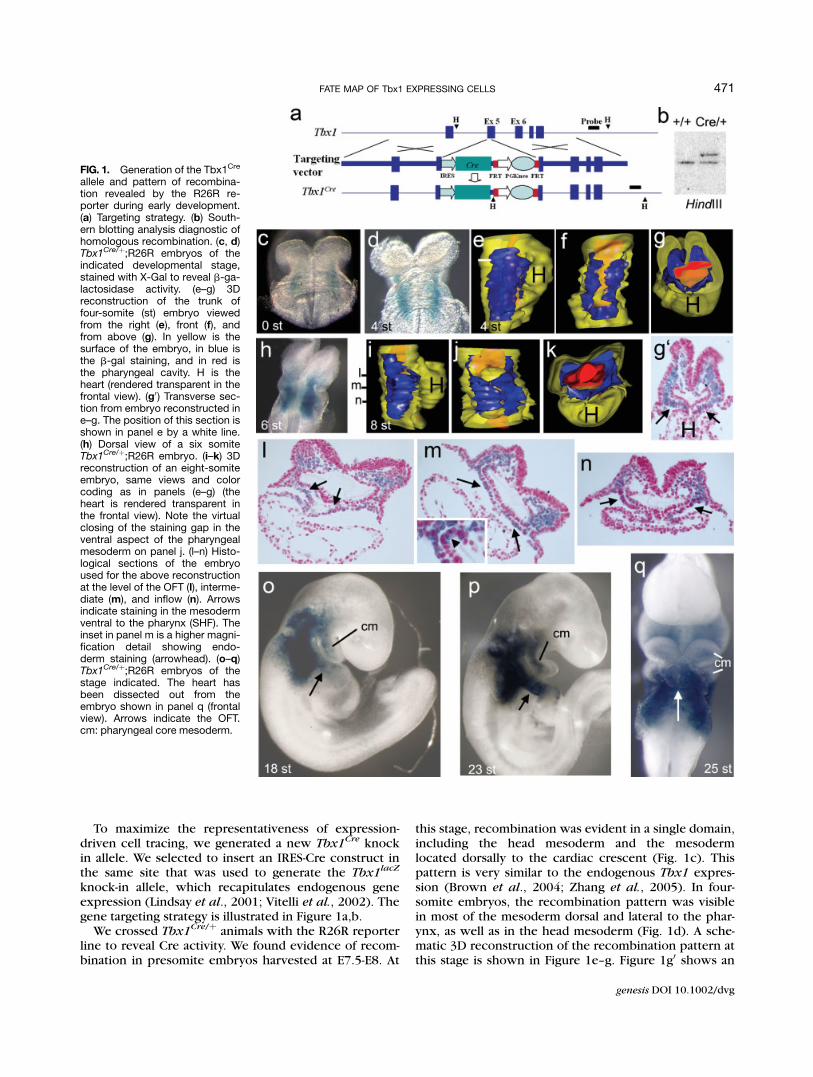
this stage, recombination was evident in a single domain,including the head mesoderm and the mesodermlocated dorsally to the cardiac crescent (Fig. 1c). Thispattern is very similar to the endogenous Tbx1 expres-sion (Brown et al., 2004; Zhang et al., 2005). In four-somite embryos, the recombination pattern was visiblein most of the mesoderm dorsal and lateral to the phar-ynx, as well as in the head mesoderm (Fig. 1d). A sche-matic 3D reconstruction of the recombination pattern atthis stage is shown in Figure 1e–g. Figure 1g0 shows an
FIG. 1. Generation of the Tbx1Cre
allele and pattern of recombina-tion revealed by the R26R re-porter during early development.(a) Targeting strategy. (b) South-ern blotting analysis diagnostic ofhomologous recombination. (c, d)Tbx1Cre/þ;R26R embryos of theindicated developmental stage,stained with X-Gal to reveal b-ga-lactosidase activity. (e–g) 3Dreconstruction of the trunk offour-somite (st) embryo viewedfrom the right (e), front (f), andfrom above (g). In yellow is thesurface of the embryo, in blue isthe b-gal staining, and in red isthe pharyngeal cavity. H is theheart (rendered transparent in thefrontal view). (g0) Transverse sec-tion from embryo reconstructed ine–g. The position of this section isshown in panel e by a white line.(h) Dorsal view of a six somiteTbx1Cre/þ;R26R embryo. (i–k) 3Dreconstruction of an eight-somiteembryo, same views and colorcoding as in panels (e–g) (theheart is rendered transparent inthe frontal view). Note the virtualclosing of the staining gap in theventral aspect of the pharyngealmesoderm on panel j. (l–n) Histo-logical sections of the embryoused for the above reconstructionat the level of the OFT (l), interme-diate (m), and inflow (n). Arrowsindicate staining in the mesodermventral to the pharynx (SHF). Theinset in panel m is a higher magni-fication detail showing endo-derm staining (arrowhead). (o–q)Tbx1Cre/þ;R26R embryos of thestage indicated. The heart hasbeen dissected out from theembryo shown in panel q (frontalview). Arrows indicate the OFT.cm: pharyngeal core mesoderm.
471FATE MAP OF Tbx1 EXPRESSING CELLS
genesis DOI 10.1002/dvg
histological section with similar orientation of Figure 1g.At the six-somite stage the pattern was very similar (Fig.1h), whereas at the eight-somite stage, the mesodermaldomain was expanded ventrally to, and thus virtuallysurrounding the pharynx (Fig. 1i–n); in addition, recom-bination was also visible in part of the pharyngeal endo-derm (arrowhead in inset of Fig. 1m).
Gene expression-based cell tracing and DiI labelingindicated that the SHF population of migratory car-diac progenitors contributes to the outflow tract ofthe heart (OFT), the right ventricle (RV), and part ofthe atria (Kelly et al., 2001; Mjaatvedt et al., 2001;Waldo et al., 2001). Some Tbx1-traced cells were al-ready detectable at the OFT region of four-somiteembryos (Fig. 1g0), but in eight-somite embryos,there was strong labeling in the mesoderm ventral tothe pharynx, as described above, (Fig. 1j,l–n arrows),including the outflow (Fig. 1l) and inflow regions(Fig. 1n). These are the domains detected by theSHF tracers Isl1
Cre, mef2c-AHF-Cre, and Mlc1v-24 atthis stage (Cai et al., 2003; Kelly et al., 2001; Verziet al., 2005). The dynamics of Tbx1
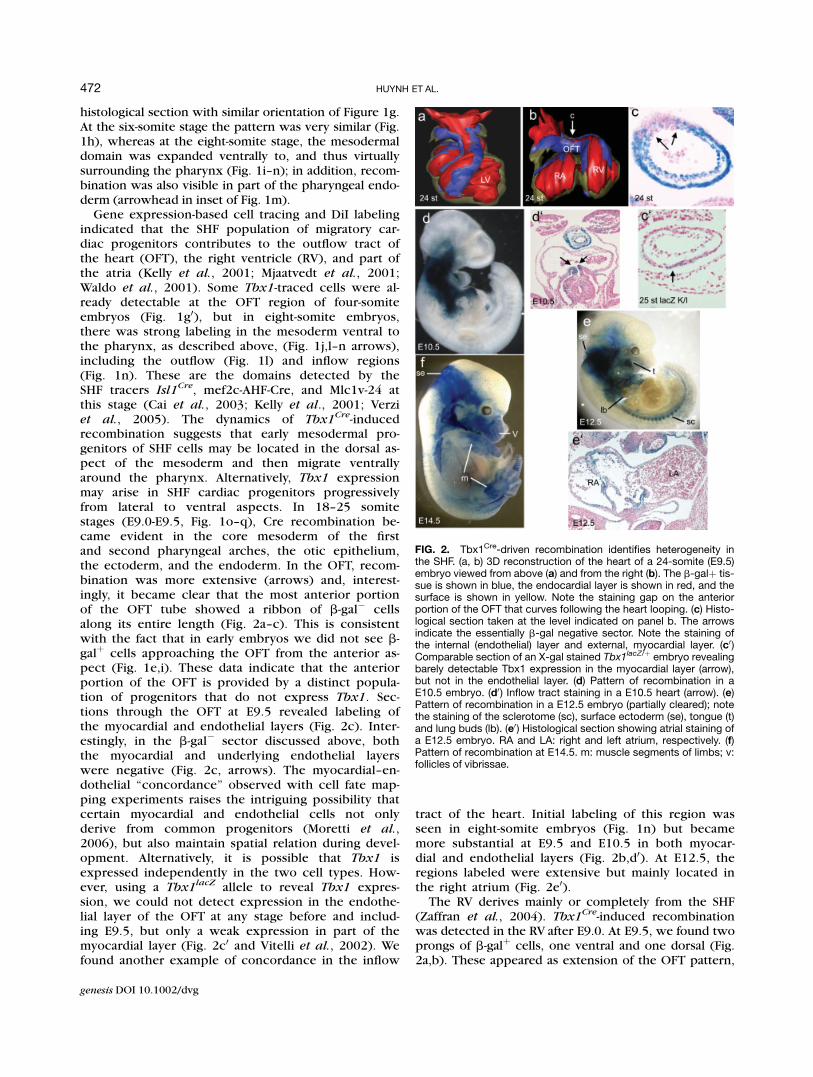
Cre-inducedrecombination suggests that early mesodermal pro-genitors of SHF cells may be located in the dorsal as-pect of the mesoderm and then migrate ventrallyaround the pharynx. Alternatively, Tbx1 expressionmay arise in SHF cardiac progenitors progressivelyfrom lateral to ventral aspects. In 18–25 somitestages (E9.0-E9.5, Fig. 1o–q), Cre recombination be-came evident in the core mesoderm of the firstand second pharyngeal arches, the otic epithelium,the ectoderm, and the endoderm. In the OFT, recom-bination was more extensive (arrows) and, interest-ingly, it became clear that the most anterior portionof the OFT tube showed a ribbon of b-gal� cellsalong its entire length (Fig. 2a–c). This is consistentwith the fact that in early embryos we did not see b-galþ cells approaching the OFT from the anterior as-pect (Fig. 1e,i). These data indicate that the anteriorportion of the OFT is provided by a distinct popula-tion of progenitors that do not express Tbx1. Sec-tions through the OFT at E9.5 revealed labeling ofthe myocardial and endothelial layers (Fig. 2c). Inter-estingly, in the b-gal� sector discussed above, boththe myocardial and underlying endothelial layerswere negative (Fig. 2c, arrows). The myocardial–en-dothelial ‘‘concordance’’ observed with cell fate map-ping experiments raises the intriguing possibility thatcertain myocardial and endothelial cells not onlyderive from common progenitors (Moretti et al.,2006), but also maintain spatial relation during devel-opment. Alternatively, it is possible that Tbx1 isexpressed independently in the two cell types. How-ever, using a Tbx1
lacZ allele to reveal Tbx1 expres-sion, we could not detect expression in the endothe-lial layer of the OFT at any stage before and includ-ing E9.5, but only a weak expression in part of themyocardial layer (Fig. 2c0 and Vitelli et al., 2002). Wefound another example of concordance in the inflow
tract of the heart. Initial labeling of this region wasseen in eight-somite embryos (Fig. 1n) but becamemore substantial at E9.5 and E10.5 in both myocar-dial and endothelial layers (Fig. 2b,d0). At E12.5, theregions labeled were extensive but mainly located inthe right atrium (Fig. 2e0).
The RV derives mainly or completely from the SHF(Zaffran et al., 2004). Tbx1
Cre-induced recombinationwas detected in the RV after E9.0. At E9.5, we found twoprongs of b-galþ cells, one ventral and one dorsal (Fig.2a,b). These appeared as extension of the OFT pattern,
FIG. 2. Tbx1Cre-driven recombination identifies heterogeneity inthe SHF. (a, b) 3D reconstruction of the heart of a 24-somite (E9.5)embryo viewed from above (a) and from the right (b). The b-galþ tis-sue is shown in blue, the endocardial layer is shown in red, and thesurface is shown in yellow. Note the staining gap on the anteriorportion of the OFT that curves following the heart looping. (c) Histo-logical section taken at the level indicated on panel b. The arrowsindicate the essentially b-gal negative sector. Note the staining ofthe internal (endothelial) layer and external, myocardial layer. (c0)Comparable section of an X-gal stained Tbx1lacZ/þ embryo revealingbarely detectable Tbx1 expression in the myocardial layer (arrow),but not in the endothelial layer. (d) Pattern of recombination in aE10.5 embryo. (d0) Inflow tract staining in a E10.5 heart (arrow). (e)Pattern of recombination in a E12.5 embryo (partially cleared); notethe staining of the sclerotome (sc), surface ectoderm (se), tongue (t)and lung buds (lb). (e0) Histological section showing atrial staining ofa E12.5 embryo. RA and LA: right and left atrium, respectively. (f)Pattern of recombination at E14.5. m: muscle segments of limbs; v:follicles of vibrissae.
472 HUYNH ET AL.
genesis DOI 10.1002/dvg
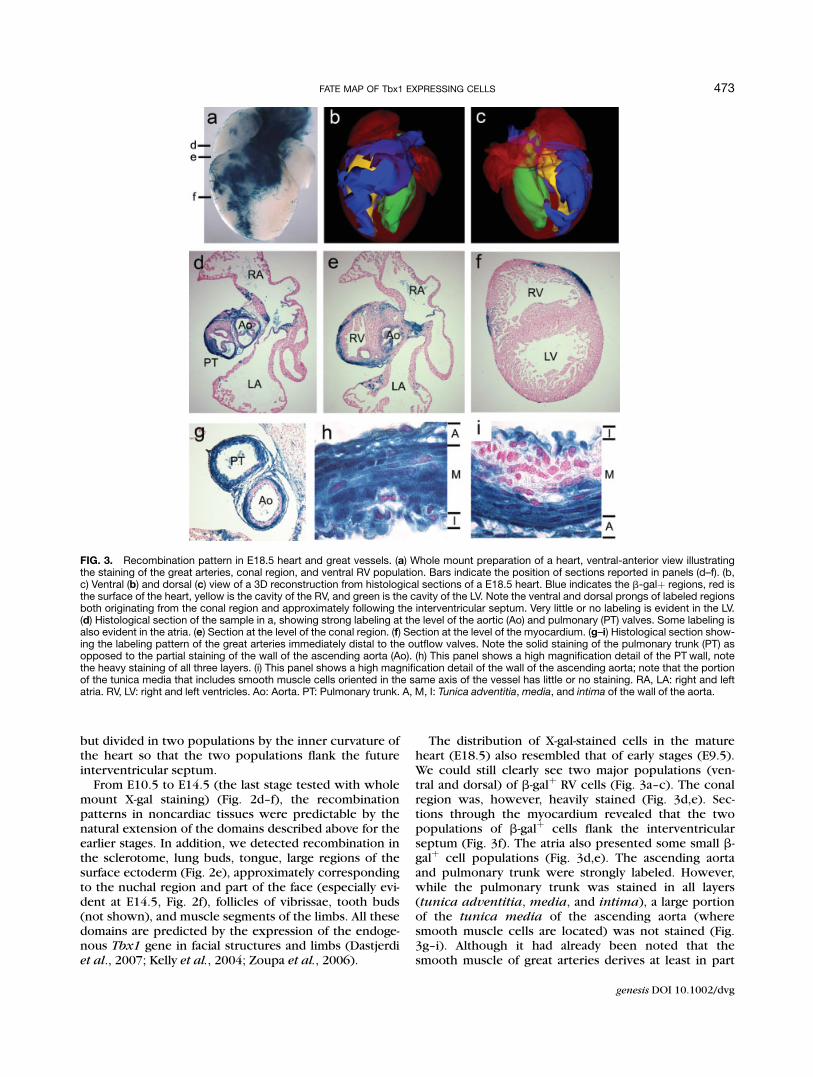
but divided in two populations by the inner curvature ofthe heart so that the two populations flank the futureinterventricular septum.
From E10.5 to E14.5 (the last stage tested with wholemount X-gal staining) (Fig. 2d–f), the recombinationpatterns in noncardiac tissues were predictable by thenatural extension of the domains described above for theearlier stages. In addition, we detected recombination inthe sclerotome, lung buds, tongue, large regions of thesurface ectoderm (Fig. 2e), approximately correspondingto the nuchal region and part of the face (especially evi-dent at E14.5, Fig. 2f), follicles of vibrissae, tooth buds(not shown), and muscle segments of the limbs. All thesedomains are predicted by the expression of the endoge-nous Tbx1 gene in facial structures and limbs (Dastjerdiet al., 2007; Kelly et al., 2004; Zoupa et al., 2006).
The distribution of X-gal-stained cells in the matureheart (E18.5) also resembled that of early stages (E9.5).We could still clearly see two major populations (ven-tral and dorsal) of b-galþ RV cells (Fig. 3a–c). The conalregion was, however, heavily stained (Fig. 3d,e). Sec-tions through the myocardium revealed that the twopopulations of b-galþ cells flank the interventricularseptum (Fig. 3f). The atria also presented some small b-galþ cell populations (Fig. 3d,e). The ascending aortaand pulmonary trunk were strongly labeled. However,while the pulmonary trunk was stained in all layers(tunica adventitia, media, and intima), a large portionof the tunica media of the ascending aorta (wheresmooth muscle cells are located) was not stained (Fig.3g–i). Although it had already been noted that thesmooth muscle of great arteries derives at least in part
FIG. 3. Recombination pattern in E18.5 heart and great vessels. (a) Whole mount preparation of a heart, ventral-anterior view illustratingthe staining of the great arteries, conal region, and ventral RV population. Bars indicate the position of sections reported in panels (d–f). (b,c) Ventral (b) and dorsal (c) view of a 3D reconstruction from histological sections of a E18.5 heart. Blue indicates the b-galþ regions, red isthe surface of the heart, yellow is the cavity of the RV, and green is the cavity of the LV. Note the ventral and dorsal prongs of labeled regionsboth originating from the conal region and approximately following the interventricular septum. Very little or no labeling is evident in the LV.(d) Histological section of the sample in a, showing strong labeling at the level of the aortic (Ao) and pulmonary (PT) valves. Some labeling isalso evident in the atria. (e) Section at the level of the conal region. (f) Section at the level of the myocardium. (g–i) Histological section show-ing the labeling pattern of the great arteries immediately distal to the outflow valves. Note the solid staining of the pulmonary trunk (PT) asopposed to the partial staining of the wall of the ascending aorta (Ao). (h) This panel shows a high magnification detail of the PT wall, notethe heavy staining of all three layers. (i) This panel shows a high magnification detail of the wall of the ascending aorta; note that the portionof the tunica media that includes smooth muscle cells oriented in the same axis of the vessel has little or no staining. RA, LA: right and leftatria. RV, LV: right and left ventricles. Ao: Aorta. PT: Pulmonary trunk. A, M, I: Tunica adventitia,media, and intima of the wall of the aorta.
473FATE MAP OF Tbx1 EXPRESSING CELLS
genesis DOI 10.1002/dvg

from the SHF (Waldo et al., 2005), our data reveal thatthere is heterogeneity in the progenitor population,even within the aorta.
In conclusion, our cell fate mapping data reveal celllineage heterogeneity within the SHF. In particular, ante-rior cells of the OFT appear to derive from a distinctcranial/medial pharyngeal population, whereas Tbx1-traced cells approach the outflow from caudal/medialand lateral directions. The Tbx1- and non-Tbx1-tracedpopulations within the OFT and RV remain distinctthroughout development, with very little or no in-termingling.
Finally, as heart-contributing progenitors appear tohave at least partially distinct transcription programs,it is reasonable to predict that mutation of differentgenes may affect different populations of otherwiseundistinguishable heart cells. Because the distributionof these cells follow a relatively rigid scheme, the lossor perturbation of a particular population will likelylead to morphogenetic abnormalities and congenitalheart disease.
METHODS
We injected into mouse blastocysts two independentlytargeted embryonic stem cell clones, both of which pro-duced germ line chimeras. Two lines were establishedand early experiments with the Cre-reporter line R26Rled to identical results. Therefore, in subsequent experi-ments, we used the two lines interchangeably. Genotyp-ing of the founders was carried out by Southern blottinganalysis (Fig. 1b); subsequent litters were genotyped byPCR using the following primers: Tbx1cre2-tar-R:50TCGCCTTCTTGACGATTCT30, Tbx1cre2-tar-F: 50GGCCTACAACAGGAGACAGC30, Tbx1cre2-wt-R: 50CGCACAGTGGATGAAACAGA30.
All the crosses were done in a C57Bl6–129SvEv mixedbackground, and we used the R26R line (Soriano, 1999)as a Cre reporter. Tbx1Cre/þ; R26R embryos were fixedin 4% paraformaldehyde and stained with X-gal O.N.according to standard procedures. At least three to fiveembryos per developmental stage were stained, photo-graphed, and then embedded in paraffin to obtain histo-logical sections. For some representative embryos, weused digital images for 3D reconstruction using the soft-ware WinSURF (provided by SurfDriver, http://www.surfdriver.com/). In particular, for embryos of four andeight somites, we used whole-embryo transverse sec-tions 7 lm-thick. Digital images of all the sections of therelevant region of the embryos (40–45 sections). For the24-somnite embryo, we used digital images of 41 consec-utive coronal sections (10 lm) that included the heart.For E18.5 hearts we used digital images of 30 transversesections (one every four consecutive histological sec-tions 10 lm thick). In all cases, digital images directlyfrom the microscope were entered into the softwareand the regions of interest in the images were identifiedmanually.
ACKNOWLEDGMENTS
We wish to acknowledge the service provided by theDarwin Transgenic Core of Baylor College of Medicine,Houston.
LITERATURE CITED
Baldini A. 2005. Dissecting contiguous gene defects: TBX1. Curr OpinGenet Dev 15:279–284.
Brown CB, Wenning JM, Lu MM, Epstein DJ, Meyers EN, Epstein JA.2004. Cre-mediated excision of Fgf8 in the Tbx1 expression do-main reveals a critical role for Fgf8 in cardiovascular developmentin the mouse. Dev Biol 267:190–202.
Cai CL, Liang X, Shi Y, Chu PH, Pfaff SL, Chen J, Evans S. 2003. Isl1 iden-tifies a cardiac progenitor population that proliferates prior to dif-ferentiation and contributes a majority of cells to the heart. DevCell 5:877–889.
Chapman DL, Garvey N, Hancock S, Alexiou M, Agulnik SI, Gibson-Brown JJ, Cebra-Thomas J, Bollag RJ, Silver LM, Papaioannou VE.1996. Expression of the T-box family genes. Tbx1–Tbx5, duringearly mouse development. Dev Dyn 206:379–390.
Dastjerdi A, Robson L, Walker R, Hadley J, Zhang Z, Rodriguez-Nieden-fuhr M, Ataliotis P, Baldini A, Scambler P, Francis-West P. 2007.Tbx1 regulation of myogenic differentiation in the limb and cranialmesoderm. Dev Dyn 236:353–363.
Kelly RG, Brown NA, Buckingham ME. 2001. The arterial pole of themouse heart forms from Fgf10-expressing cells in pharyngeal mes-oderm. Dev Cell 1:435–440.
Kelly RG, Jerome-Majewska LA, Papaioannou VE. 2004. The del22q11.2candidate gene Tbx1 regulates branchiomeric myogenesis. HumMol Genet 13:2829–2840.
Kelly RG, Papaioannou VE. 2007. Visualization of outflow tract develop-ment in the absence of Tbx1 using an FgF10 enhancer trap trans-gene. Dev Dyn 236:821–828.
Lindsay EA, Vitelli F, Su H, Morishima M, Huynh T, Pramparo T,Jurecic V, Ogunrinu G, Sutherland HF, Scambler PJ, Bradley A,Baldini A. 2001. Tbx1 haploinsufficieny in the DiGeorge syn-drome region causes aortic arch defects in mice. Nature410:97–101.
Maeda J, Yamagishi H, McAnally J, Yamagishi C, Srivastava D. 2006.Tbx1 is regulated by forkhead proteins in the secondary heartfield. Dev Dyn 235:701–710.
Mjaatvedt CH, Nakaoka T, Moreno-Rodriguez R, Norris RA, Kern MJ,Eisenberg CA, Turner D, Markwald RR. 2001. The outflow tract ofthe heart is recruited from a novel heart-forming field. Dev Biol238:97–109.
Moretti A, Caron L, Nakano A, Lam JT, Bernshausen A, Chen Y, QyangY, Bu L, Sasaki M, Martin-Puig S, Sun Y, Evans SM, Laugwitz KL,Chien KR. 2006. Multipotent embryonic isl1 þ progenitor cellslead to cardiac, smooth muscle, and endothelial cell diversifica-tion. Cell 127:1151–1165.
Naiche LA, Harrelson Z, Kelly RG, Papaioannou VE. 2005. T-boxgenes in vertebrate development. Annu Rev Genet 39:219–239.
Paylor R, Glaser B, Mupo A, Ataliotis P, Spencer C, Sobotka A, Sparks C,Choi CH, Oghalai J, Curran S, Murphy KC, Monks S, Williams N,O’Donovan MC, Owen MJ, Scambler PJ, Lindsay E. 2006. Tbx1haploinsufficiency is linked to behavioral disorders in mice andhumans: Implications for 22q11 deletion syndrome. Proc NatlAcad Sci U S A 103:7729–7734.
Soriano P. 1999. Generalized lacZ expression with the ROSA26 Cre re-porter strain [letter]. Nat Genet 21:70–71.
Verzi MP, McCulley DJ, De Val S, Dodou E, Black BL. 2005. The rightventricle, outflow tract, and ventricular septum comprise a re-stricted expression domain within the secondary/anterior heartfield. Dev Biol 287:134–145.
Vitelli F, Morishima M, Taddei I, Lindsay EA, Baldini A. 2002. Tbx1muta-tion causes multiple cardiovascular defects and disrupts neuralcrest and cranial nerve migratory pathways. Hum Mol Genet11:915–922.
474 HUYNH ET AL.
genesis DOI 10.1002/dvg

Waldo KL, Hutson MR, Ward CC, Zdanowicz M, Stadt HA, Kumiski D,Abu-Issa R, Kirby ML. 2005. Secondary heart field contributes myo-cardium and smooth muscle to the arterial pole of the developingheart. Dev Biol 281:78–90.
Waldo KL, Kumiski DH, Wallis KT, Stadt HA, Hutson MR, Platt DH,Kirby ML. 2001. Conotruncal myocardium arises from a secondaryheart field. Development 128:3179–3188.
Xu H, Cerrato F, Baldini A. 2005. Timed mutation and cell-fate mappingreveal reiterated roles of Tbx1 during embryogenesis, and a crucialfunction during segmentation of the pharyngeal system via regula-tion of endoderm expansion. Development 132:4387–4395.
Xu H, Morishima M, Wylie JN, Schwartz RJ, Bruneau BG, Lindsay EA,Baldini A. 2004. Tbx1 has a dual role in the morphogenesis of thecardiac outflow tract. Development 131:3217–3227.
Xu H, Viola A, Zhang Z, Gerken CP, Lindsay-Illingworth EA, Baldini A.2007. Tbx1 regulates population, proliferation and cell fate deter-mination of otic epithelial cells. Developmental Biology 302:670–682.
Yagi H, Furutani Y, Hamada H, Sasaki T, Asakawa S, Minoshima S, IchidaF, Joo K, Kimura M, Imamura S, Kamatani N, Momma K, Takao A,Nakazawa M, Shimizu N, Matsuoka R. 2003. Role of TBX1 inhuman del22q11.2 syndrome. Lancet 362:1366–1373.
Zaffran S, Kelly RG, Meilhac SM, Buckingham ME, Brown NA. 2004.Right ventricular myocardium derives from the anterior heart field.Circ Res 95:261–268.
Zhang Z, Cerrato F, Xu H, Vitelli F, Morishima M, Vincentz J, Furuta Y,Ma L, Martin JF, Baldini A, Lindsay E. 2005. Tbx1 expression in pha-ryngeal epithelia is necessary for pharyngeal arch artery develop-ment. Development 132:5307–5315.
Zhang Z, Huynh T, Baldini A. 2006. Mesodermal expression of Tbx1 isnecessary and sufficient for pharyngeal arch and cardiac outflowtract development. Development 133:3587–3595.
Zoupa M, Seppala M, Mitsiadis T, Cobourne MT. 2006. Tbx1 isexpressed at multiple sites of epithelial–mesenchymal interactionduring early development of the facial complex. Int J Dev Biol50:504–510.
475FATE MAP OF Tbx1 EXPRESSING CELLS
genesis DOI 10.1002/dvg