Embed Size (px)
Citation preview

The Mechanism of Iodide Accumulation by the Brown Sea Weed Laminaria digitata. II.Respiration and Iodide UptakeAuthor(s): T. I. ShawSource: Proceedings of the Royal Society of London. Series B, Biological Sciences, Vol. 152, No.946, A Discussion on the 'Ear' Under Water (Apr. 26, 1960), pp. 109-117Published by: The Royal SocietyStable URL: http://www.jstor.org/stable/75367 .
Accessed: 08/05/2014 12:19
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The Royal Society is collaborating with JSTOR to digitize, preserve and extend access to Proceedings of theRoyal Society of London. Series B, Biological Sciences.
http://www.jstor.org
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 12:19:20 PMAll use subject to JSTOR Terms and Conditions

The mechanism of iodide accumulation by the brown sea weed Laminaria digitata
II. Respiration and iodide uptake
By T. I. SHAW
The Mlarine Biological Laboratory, Plymouth
(Communicated by F. S. Russell, F.R.S.-Received 6 August 1959)
Iodide absorption by fresh, summer Laminaria digitata frond was accompanied by about a fivefold increase in the rate of oxygen consumption. Between approximately 3 and 6 mole- cules of O0 were consumed for each iodide ion absorbed by the tissue and the respiratory quotient, whether measured in the presence or absence of iodide, lay between 0-9 and 1.1 (mean 0-94).
It is argued that: (1) The iodide-stimulated respiration must occur principally near the surface of the frond,
though this localization is not due to the limited diffusibility of oxygen. (2) Iodide absorption is connected with the oxidation of a carbohydrate-like compound or
compounds. (3) The absorption of iodide is associated with a rate-limiting process for the oxidation of
the compound or compounds. (4) Between one and two iodide ions are absorbed for each six-carbon unit oxidized. (5) Probably only one or two reactions in the oxidation of each six-carbon unit is involved
in iodide absorption, one of these reactions being the rate limiting process governing the oxidation.
INTRODUCTION
Lundegardh (1954) and Suteliffe ( 959) have reviewed much of the extensive litera- ture dealing with the close relationship between anion accumulation and respira- tion in plants. Robertson (I941) and Robertson & Wilkins (1948) have pointed out the significance that may attach to the number of ions transported for each 02
molecule consumed. In Laminaria digitata frond iodide accumulation seems to involve iodine as an oxygen carrier (Shaw I959) so that concurrent measurements of 02 utilization and iodine uptake are clearly of peculiar interest for this system.
Kelly (1953) and Klemperer (I957) have studied the effects of metabolic inhi- bitors upon both respiratory exchanges and radio-iodine uptake by brown sea weeds. Their results show that with different inhibitors the respiration may be either depressed, unaffected or even stimulated when the radio-iodine uptake is
depressed.
METHODS
With a few exceptions, noted later, Laminaria digitata plants were used within 24 h of collection from the rocks near the Plymouth laboratory.
Oxygen consumption was followed by means of a Warburg manometer. Disks of frond 0-5 cm diameter were stamped out by cork borer and washed in sea water. Generally ten disks, forming a total of roughly 180 mg fresh weight, were
placed in the main compartment of each flask together with 2-4 ml. filtered sea water. Into the central well was put 0-2 ml. 20 % KOH together with a roll of
[ 109 ]
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 12:19:20 PMAll use subject to JSTOR Terms and Conditions

filter-paper. The side arm contained 0-4 ml. sea water in which had been dissolved sodium or potassium iodide.
The experiments were carried out in the dark at 19-5 ?C with a stroke frequency of 100 min-1; increasing the stroke frequency did not affect the 02 consumption whether measured in the presence or absence of iodide. Readings were taken about every 45 min. Generally air, though occasionally O2 at atmospheric pressure, formed the ambient gas. The procedure allowed 5 min for the flasks to attain bath
temperature before the joints, sealed with Vaseline, were ground in. After a further 15 min the tap to the atmosphere was closed and measurements started. The basal respiration was followed over 2 to 4 h before the flask was tipped to introduce iodide into the main compartment.
The total extra O2 consumption due to iodide was determined by plotting the results, extrapolating back the final linear portion of the curve (see figure 1), and
measuring the displacement between this line and the initial basal 02 consumption at the time of addition of iodide. In the construction of figure 2, where the time course of the extra oxygen consumption has been plotted, the initial linear portion of the curve was extrapolated and the displacement of subsequent measurements from this line plotted.
Objection might be raised to the use of sea water as a suspending medium in
Warburg flasks since loss of CO2 to the KOH of the central well might be expected to leave a highly alkaline solution. Surprisingly it was found that, in the presence of weed, the suspending water, originally pH 8-1, tended to become more acid
(pH 7-7 to 7-9 by glass electrode). This may be caused by the evasion of CO2 from the sea water being slower than its production by the tissue.
Further objection might be raised to the experiments since iodide is oxidized to iodine by L. digitata and the free I2 can be expected to diffuse into the alkali of the central well causing a loss other than by absorption into the weed. That this is not serious has been shown by the addition of 131 to a flask containing sea water, weed, KI and the usual alkali and filter-paper in the central well. After 1.5 h less than 2 % of the activity originally added to the flask was found in the KOH and filter-
paper; this small loss has been neglected in the subsequent account and discussion of the results.
Respiratory quotients were determined by the direct method, described by Dixon (1934) and involved three flasks for each estimation. CO2 was driven from sea water and tissues by 3 N-HCI, of which two flasks from each triplet contained 0-4 ml. in their side arms.
To convert manometer readings to gas volumes the manometers were calibrated with Hg and their constants calculated from the data quoted by Harvey (1955) for the solubilities of 02 and CO2 in sea water.
Iodide was detected by the addition of HC1, starch and nitrite solutions; estima- tion was carried out with only slight modification of this procedure, the optical densities of the blue colour at 615 m/u being compared with those obtained using standard iodide solutions identically treated. Corresponding values of iodide
absorption and O2 consumption (as given in figure 2) were obtained using several
Warburg flasks containing samples of weed from the same plant and maintained
110 T. I. Shaw
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 12:19:20 PMAll use subject to JSTOR Terms and Conditions

Iodide and respiration in Laminaria digitata. II 111
under identical conditions. The 02 consumption was followed in one flask, while the other vessels were opened at various times and the unabsorbed iodine estimated as described above.
EXPERIMENTAL RESULTS
The slow and steady 02 consumption which occurs when weed is kept in the absence of iodide and in the dark is shown by the early portions of the curves in
figure 1, and data concerning this basal rate are summarized in the upper part of table 1. Replacement of air by 02 as the manometric gas failed to cause marked enhancement of the 02 uptake (table 2).
TABLE 1. THE RATE OF 02 CONSUMPTION BY LAMINARIA DIGITATA FROND
IN THE PRESENCE AND ABSENCE OF ADDED IODIDE
no. of 1A. 02 no. of / 1. 02 no. of /1. 02 no. of disks of cm-2 h-l experi- (g fresh)-1 h-l experi- (mg dry)-1 h-1 experi-
iodide weed/flask + s.d. ments + s.d. ments ? s.d. ments
- 10 8-4+ 1-7 31 86 + 17 21 0-64 ?+ 014 21 - 20 8-3 1 86 1 0-61 1 - 20 8.0 1 80 1 0.60 1 - 20 7.6 1 ---
+ 10 40 ?13 26 416* ?85 18 3.1 ?+07 14
+ 20 29 1 283 1 2-1 1
+ 20 49 1 510 1 3-6 1
Manometric gas, air; suspending medium, sea water. * Mean tissue thickness 8 9 x 10-2 cm, mean (tissue thickness)2 = 8.1 x 10-3 cm2 in these experiments. Tissue density 1-06 g ml.-'
(Shaw 1959).
TABLE 2. RELATIVE RATES OF 02 CONSUMPTION BY LAMINVARIIA DIGITATA FROND
IN AIR AND IN 02 no. of
rate in air relative rate in paired iodide taken as 02 + s.e.m. samples
-- 1.0 1.13 + 0-08 8 + 1-0 1-05 + 0-08 8
The rate (per unit fresh weight) in 02 is stated in relation to the rate found for tissue from the same plant in air.
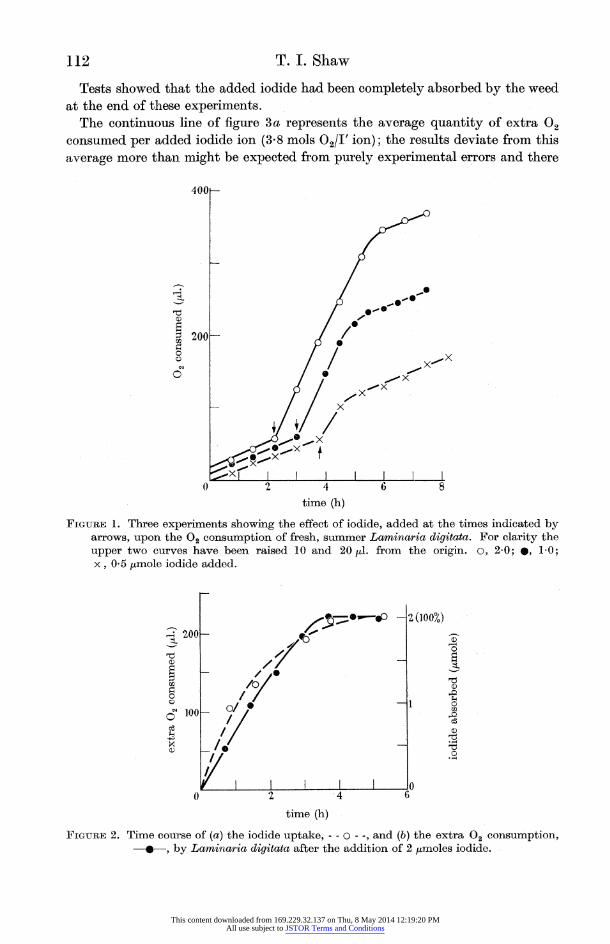
When iodide was added to fresh, summer weed there was a marked and sudden increase of oxygen consumption (figure 1). The increase was only temporary and the oxygen consumption eventually returned to a value close to that prevailing before the addition of iodide.
The end of the respiratory burst approximately coincided with the completion of iodine absorption by the weed. This is shown in figure 2 where the extra oxygen consumption due to added iodide and the iodide absorbed by the weed are plotted on the same graph.
The quantity of extra oxygen consumed was directly related to the amount of iodide absorbed by the weed. This is shown by the three curves illustrated in
figure 1 which correspond to the addition of three different amounts of added
iodide, and by the collected data shown in figure 3a.
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 12:19:20 PMAll use subject to JSTOR Terms and Conditions
112 T. I. Shaw
Tests showed that the added iodide had been completely absorbed by the weed at the end of these experiments.
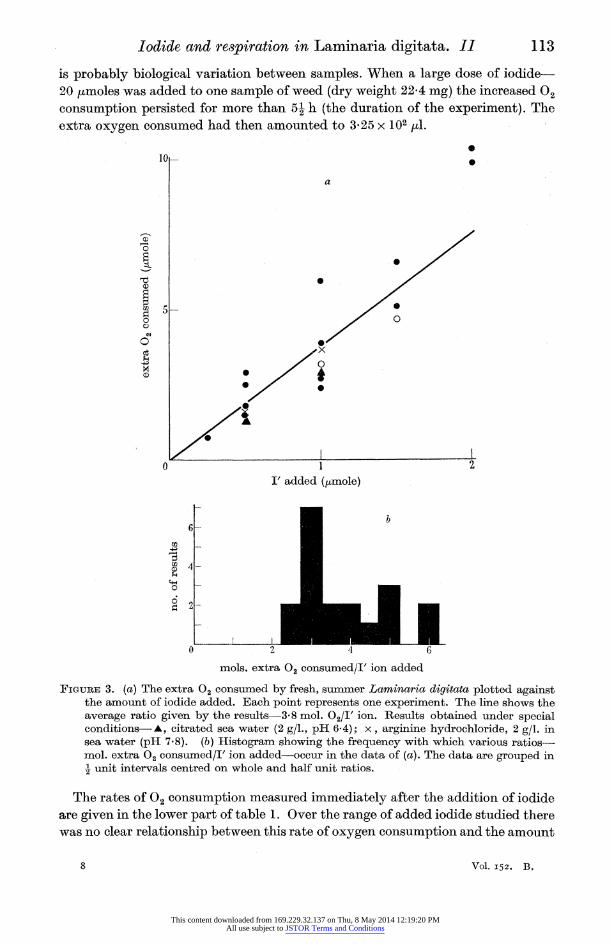
The continuous line of figure 3a represents the average quantity of extra 02 consumed per added iodide ion (3-8 mols O2/I' ion); the results deviate from this
average more than might be expected from purely experimental errors and there
400-
200- / O
Y.:(/
0 2 4 6 8 time (h)
FIGURE 1. Three experiments showing the effect of iodide, added at the times indicated by arrows, upon the 02 consumption of fresh, summer Laminaria digitata. For clarity the upper two curves have been raised 10 and 20 /d. from the origin. o, 2-0; *, 1-0; x, 0-5 /mole iodide added.
4 200
It
o 0
o 100 0
C !
0
2 (100%)
1 I
a>
0
0
0 0..
? -
_ o/? 0 // 0
/- / -
/II
I,,I
2 4 6 n
time (h)
FIGUTRE 2. Time course of (a) the iodide uptake, - - O - -, and (b) the extra 02 consumption, ---, by Laminaria digitata after the addition of 2 ,Imoles iodide.
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 12:19:20 PMAll use subject to JSTOR Terms and Conditions
Iodide and respiration in Laminaria digitata. II 113
is probably biological variation between samples. When a large dose of iodide- 20 /moles was added to one sample of weed (dry weight 22-4 mg) the increased 02
consumption persisted for more than 5- h (the duration of the experiment). The extra oxygen consumed had then amounted to 3.25 x 102 pi.
10-
a
(3) 0
i
1-p
0
(1)
5
0
0 0
0
A/! 0.
added (mo) I' added (txmole)
Z 4
mols. extra 02 consumed/I' ion added
FIGURE 3. (a) The extra 02 consumed by fresh, summer Laminaria digitata plotted against the amount of iodide added. Each point represents one experiment. The line shows the average ratio given by the results-3 8 mol. 02/I' ion. Results obtained under special conditions--A, citrated sea water (2 g/1., pHI 6.4); x, arginine hydrochloride, 2 g/l. in sea water (pH 7.8). (b) Histogram showing the frequency with which various ratios- mol. extra 02 consumed/I' ion added-occur in the data of (a). The data are grouped in -I unit intervals centred on whole and half unit ratios.
The rates of 02 consumption measured immediately after the addition of iodide are given in the lower part of table 1. Over the range of added iodide studied there was no clear relationship between this rate of oxygen consumption and the amount
Vol. 152. B.
6
4
rh
4.1 0 ?
Ft
I 6
0
_ -I ,r I
8
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 12:19:20 PMAll use subject to JSTOR Terms and Conditions
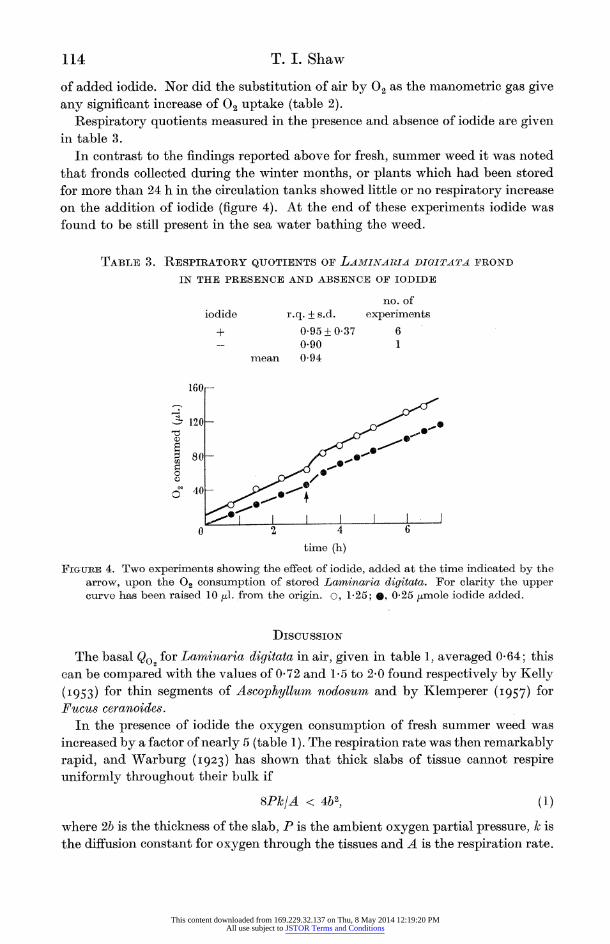
of added iodide. Nor did the substitution of air by 02 as the manometric gas give any significant increase of O2 uptake (table 2).
Respiratory quotients measured in the presence and absence of iodide are given in table 3.
In contrast to the findings reported above for fresh, summer weed it was noted that fronds collected during the winter months, or plants which had been stored for more than 24 h in the circulation tanks showed little or no respiratory increase on the addition of iodide (figure 4). At the end of these experiments iodide was found to be still present in the sea water bathing the weed.
TABLE 3. RESPIRATORY QUOTIENTS OF LAMLINARInAI DIGITAT1A FROND
IN THE PRESENCE AND ABSENCE OF IODIDE
no. of iodide r.q. ? s.d. experiments
+ 0.95+0.37 6 0-90 1
mean 0-94
160-
120- .e
1 80- '"'
040-
&1 I I. 1 1 0 0 2 4 6
time (h)
FIGURE 4. Two experiments showing the effect of iodide, added at the time indicated by the arrow, upon the 02 consumption of stored Laminaria digitata. For clarity the upper curve has been raised 10 tul. from the origin. o, 1-25; 9, 0.25 ttmole iodide added.
DISCUSSION
The basal Qo02 for Laminaria digitata in air, given in table 1, averaged 0-64; this can be compared with the values of 0-72 and 1-5 to 2-0 found respectively by Kelly (1953) for thin segments of Ascophyllum nodosum and by Klemperer (I957) for Fucus ceranoides.
In the presence of iodide the oxygen consumption of fresh summer weed was increased by a factor of nearly 5 (table 1). The respiration rate was then remarkably rapid, and Warburg (I923) has shown that thick slabs of tissue cannot respire uniformly throughout their bulk if
8Pk/A < 4b2, (1)
where 2b is the thickness of the slab, P is the ambient oxygen partial pressure, k is the diffusion constant for oxygen through the tissues and A is the respiration rate.
114 T. I. Shaw
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 12:19:20 PMAll use subject to JSTOR Terms and Conditions

Iodide and respiration in Laminaria digitata. II
The mean values from table 1 relating to experiments with air (P = 0-21 atm) are 4b2 = 8-1 x 10-3 cm2 and A == 7-35 x 10-3 ml. in- ml.-l. The diffusion constant ik has not been determined for Laminaria frond: for 20 ?C Krogh (I919) gives its value in animal connective tissue as 1-15 x 10-5 cm2 min-l atm-1 and quotes the
figure of 3.4 x 10-5 for water. Using the former figure for k the left-hand side of (1) takes the value of 2-6 x 10-3, using the latter it takes the value 7.8 x 10-3; in both cases 8 Pk/A < 4b2. Therefore (unless protoplasmic streaming hastens the dis- semination of O2 through the frond) the oxygen consumption in the presence of iodide must occur predominantly towards the surface of the plant. Such localiza- tion might be due to the relatively slow diffusion of oxygen through the tissues, but if this were so an increase of 02 partial pressure, by hastening diffusion and ex-
posing more tissue to that gas, should enhance the oxygen consumption. Table 2 shows that no such effect occurred. The localization of metabolic machinery, or the site of iodine absorption, presumably determine the siting of the oxygen consump- tion towards the weed's surface.
The substrate or substrates oxidized during iodine uptake appeared to be mainly carbohydrates or carbohydrate-like compounds since the respiratory quotient (r.q.) is close to unity (table 3). An extra oxygen consumption amounting overall to 1-45 x 104 1J. 02/g dry wt, observed when excess iodide was added to one plant, implied that, with substrates of the type (CnH2nO), the quantity of material oxidized amounted to 19*4 mg/g, or 1-94% of the dry weight of tissue. These
figures indicate that the compound or compounds oxidized during the iodine uptake included a major tissue component of the carbohydrate type. Black (I953) reports the principal carbohydrate-like compounds in Laminaria as mannitol (r.q. calculated = 0.923), laminarin, fucoidin, alginic acid and cellulose of which the first two are regarded as being carbohydrate reserves.
There are strong grounds for believing that sugar oxidation by biological materials is not a single stage process but involves a series of sequential reactions, the theory of which (Burton 1936) indicates that, in general, the overall rate is determined by the speed of one rate-limiting or 'master' reaction. (Restrictive conditions to the theory of the 'master' reaction are also considered by Burton
(1936) and by Hinshelwood (I946) and Krebs (I957).) Since the iodine uptake in fresh summer weed was associated with a large increase in the rate of sugar oxidation the uptake, it seems, must at least have been associated with a rate-
limiting reaction for one pathway of sugar oxidation.
Klemperer (i957) detected no respiratory increase on adding 10-3 M-iodide to Fucus ceranoides, likewise in the present experiments stored or winter weed failed to give a detectably increased oxygen consumption on the addition of iodide. It seems possible that the rate-limiting reaction for the relevant pathway for sugar oxidation is not always the same reaction and not always the reaction connected with iodine uptake, but that its identity is determined by the physiological state and perhaps by the species of plant. On the other hand, lack of suitable substrate
may determine the behaviour of stored and winter weed. It has been shown (figure 3a) that an average of 3-8 molecules of O2 were consumed
for each iodide ion added to, and absorbed by, fresh, summer weed. The same 8-2
115
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 12:19:20 PMAll use subject to JSTOR Terms and Conditions

results plotted as a histogram in figure 3 b show that in general 3, though occasion-
ally up to 6, molecules of 02 were used for each iodide ion taken up. In calculating these figures it has been assumed that the basal respiration persists during iodide
absorption. However, since the total respiration in the presence of iodide is about five times as large as the basal respiration the assumption, even if false, can only introduce errors of some 20 %.
As with iodine uptake by Fucus ceranoides (Klemperer I957) the process of iodine accumulation seems to be energetically very inefficient when related to the observed respiration. If the iodine were concentrated 30000-fold (Shaw i959) the
work-neglecting any electrical component-(Keynes & Maisel i954) would only demand the expenditure of 6000 cal, the oxidation of 0.0089 mole of glucose or, equivalently, the consumption of 0.053 mole of 02 for each g ion of iodide absorbed. Three molecules of oxygen per iodide ion are in excess of the stoicheiometric
requirement for the conversion of iodide to any recorded oxidation state as an
inorganic compound and grossly in excess of the ratio ( mol. 02/I' ion) demanded
by an efficient accumulation system connected with electron transport as con- sidered, quantitatively, by Robertson & Wilkins (I948). Remembering, however, that iodide absorption was apparently associated with sugar oxidation it is worth
noting that 3 molecules of 02 represent the quantity required to oxidize half, and 6 molecules of 02 the amount required to oxidize a whole six-carbon sugar mole- cule. The observed relationship would occur if only one, or at the most two, reactions in the breakdown of each six-carbon sugar were involved in the uptake of each iodide ion, one of these reactions being the rate limiting reaction for the
sugar's oxidation. If this were so then iodine uptake should be obligatorily related to only part of the observed extra 02 consumption. This is borne out to some extent
by the data in figure 2 where it is seen that the iodide absorption tends to precede the 02 uptake, so that the ratio mols. 02/I' ion absorbed is lower in the early stages of the experiment, than the value finally obtained. Moreover, Kelly (1953), after
studying the effects of metabolic inhibitors on respiration and 131I uptake by Ascophyllum nodosum, concluded that the iodine uptake was an aerobic process but that only a fraction of aerobic metabolism was involved in the uptake.
Evidence has been given elsewhere (Shaw I959) that iodine probably enters the weed as HIO and is largely reduced in the tissues to iodide, the energy required for absorption being expended in reducing HIO. In the present paper it has been shown that in fresh, summer weed the rate limiting reaction of one of the pathways for sugar oxidation is apparently associated with iodine absorption. It remains for further work to attempt identification of that reaction and indicate whether HIO acts as hydrogen acceptor therein.
I am grateful to Mr L. G. Hummerstone who has given valuable technical assistance throughout this work, which has been supported by the United King- dom Atomic Energy Authority.
116 T. I. Shaw
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 12:19:20 PMAll use subject to JSTOR Terms and Conditions

Iodide and respiration in Laminaria digitata. II 117
REFERENCES
Black, W. A. P. 1953 Rep. Progr. Chem. 50, 322. Burton, A. C. I936 J. Cell. Comp. Physiol. 9, 1. Dixon, M. 1934 Manometric methods, p. 45. Cambridge University Press. Harvey, H. W. 1955 The chemistry and fertility of sea waters, pp. 168, 184. Cambridge
University Press. Hinshelwood, C. N. 1946 The chemical kinetics of the bacterial cell, pp. 84 et seq. Oxford:
Clarendon Press. Kelly, S. 1953 Biol. Bull., Woods Hole, 104, 138. Keynes, R. D. & Maisel, G. W. 1954 Proc. Roy. Soc. B, 142, 383. Klemperer, H. G. I957 Biochem. J. 67, 381. Krebs, H. A. I957 Endeavour, 16, 125. Krogh, A. 19I9 J. Physiol. 52, 391.
Lundegardh, H. I954 Symp. Soc. Exp. Biol. no. 8, p. 262. Cambridge University Press. Robertson, R. N. I94I Aust. J. exp. Biol. med. Sci. 19, 265. Robertson, R. N. & Wilkins, M. 1948 Nature, Lond. 161, 101. Shaw, T. I. I959 Proc. Roy. Soc. B, 150, 356. Sutcliffe, J. F. 1959 Biol. Rev. 34, 159. Warburg, 0. I923 Biochem. Z. 142, 317.
This content downloaded from 169.229.32.137 on Thu, 8 May 2014 12:19:20 PMAll use subject to JSTOR Terms and Conditions





























