-
8/17/2019 9 Membranas Biológicas y Transporte
1/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
1 1
3. MEMBRANAS BIOLÓGICAS Y TRANSPORTE
Resulta difícil explicar la estructura y función de células y
organelos celulares sin considerar elpapel que desempeña la
membrana celular. Para llevar a cabo las reacciones químicas
necesarias en elmantenimiento de la vida, la célula necesita
mantener un medio interno apropiado. Esto es posible porquelas
células se encuentran separadas del mundo exterior por una membrana
limitante: la membranaplasmática. Además, la presencia de membranas
internas en las células eucariotas proporcionacompartimientos
adicionales que limitan ambientes únicos en los que se llevan al
cabo funcionesaltamente específicas, necesarias para la
supervivencia celular. Sin la existencia de las membranas
habríasido imposible que la vida en la tierra evolucionara hasta
alcanzar su estado actual.
Las membranas celulares no son paredes rígidas, sino estructuras
complejas y dinámicas compuestas por moléculas que poseen
características especiales. Tales características hacen posible
laexistencia de interacciones selectivas entre los sistemas de
membrana internos en la célula, y de la célula
con el medio que la rodea. Entre otras funciones de la membrana
celular, se destacan la regulación deltransporte de moléculas hacia
adentro y afuera de la célula, la transmisión de señales e
información entreel medio y el interior de la célula, la capacidad
de actuar como sistema de transferencia y almacenamientode energía
y el reconocimiento de la célula con su entorno.
Con el fin de entender los mecanismos mediante los cuales las
membranas celulares realizandichas funciones, es necesario hacer
una revisión de los conocimientos acerca de la estructura
ycomposición de las membranas en general. Aunque este tema se
centra principalmente en la estructura yfunción de la membrana
plasmática, gran parte de los conceptos que se vierten también se
aplican alsistema de membranas interno.
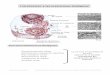
Organización de las membranas biológicas.Una de las cosas más
sorprendentes
que se descubren al observar una micrografíaelectrónica para
comparar los tamaños de lasdistintas estructuras celulares, es el
hecho deque la membrana plasmática parece serexcesivamente delgada.
Efectivamente, lamembrana plasmática tiene un grosor nomayor de 5
nm (El nanómetro es la unidad delongitud que equivale a una
milmillonésimaparte de un metro. La abreviatura del
nanómetro es nm. (1 nm = 1x10-9
m)Mucho antes de que se inventara elmicroscopio
electrónico ya se sabía que lasmembranas estaban compuestas de
proteínasy lípidos. Ya entre 1920 y 1930 se aceptabaque la parte
central de la membranaplasmática estaba formada de
lípidos,principalmente fosfolípidos. Además, el estudio de la
membrana de los eritrocitos (que sólo tienenmembrana plasmática) y
la comparación del área de superficie de membrana con el número
total demoléculas de lípidos por célula permitió a los
investigadores arribar a la conclusión de que la membrana secompone
de fosfolípidos, y que su grosor no es mayor que el de dos
moléculas de éstos. Debido a que la
mayor parte de las proteínas tiene un diámetro mayor a 10 nm,
uno de los principales problemas paracomprender la estructura
básica de las membranas consistía en determinar la forma en que las
moléculasse disponían en un espacio tan pequeño.
-
8/17/2019 9 Membranas Biológicas y Transporte
2/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
2 2
En 1972, S.T. Singer y G.L. Nicholson1 propusieron un
modelo de estructura de membranas quesintetizaba las propiedades
conocidas de las membranas biológicas. Según este modelo del
mosaicofluido1, que ha tenido gran aceptación, las membranas
constan de una bicapa lipídica (esencialmente
una doble capa de fosfolípidos) en la cual están inmersas
diversas proteínas.Esta bicapa lipídica constituye la estructura
básica de la membrana y actúa de barrerarelativamente impermeable
al paso de la mayoría de las moléculas hidrosolubles. Las moléculas
proteicas,que normalmente se hallan “disueltas” en la bicapa
lipídica, actúan como mediadores o facilitadores decasi todas las
funciones de la membrana, ya sea transportando moléculas
específicas a través de ella ocatalizando reacciones asociadas a la
membrana, como la síntesis de ATP. Algunas proteínas actúancomo
eslabones estructurales que relacionan la membrana plasmática al
citoesqueleto y/o con la matrizextracelular de las células
adyacentes, mientras que otras proteínas actúan como receptores que
reciben ytransducen las señales químicas procedentes del entorno
celular.
En la actualidad se sabe que los lípidos de membrana tienen
propiedades especiales que lespermiten formar estructuras de doble
capa, y que estas estructuras permiten la integración de
membranas
biológicas. Pero, ¿Cómo es posible que los lípidos se comporten
en esa forma?
Bicapa lipídicaLos lípidos son insolubles en agua pero se
disuelven fácilmente en disolventes orgánicos.
Constituyen aproximadamente el 50% de la masa de la mayoría de
las membranas plasmáticas de lascélulas animales, siendo casi todo
el resto proteínas. Existen 109 moléculas lipídicas en la
membranaplasmática de una célula animal pequeña.Las propiedades
físicas de los fosfolípidos, en particular la forma en que dichas
moléculas se asocian enel agua, son las que permiten laformación de
capas dobles. Ya seha mencionado que los
fosfolípidos están formados pordos cadenas de ácidos
grasosunidas a dos de los tres carbonosde la molécula del glicerol.
Lasdos cadenas de ácidos grasos dela molécula son hidrófobas
(noafines al agua) y pueden tenerdiferente longitud
(usualmentecontienen de 14 a 24 átomos decarbono). Normalmente una
deestas cadenas presenta uno omás dobles enlaces cis (es
decir,es insaturada) mientras que la otranormalmente no tiene
doblesenlaces (es saturada). Las diferencias en longitud y grado de
instauración entre las colas hidrocarbonadasson importantes porque
afectan la capacidad de las moléculas de fosfolípidos para
empaquetarse,modificando su fluidez (como veremos más adelante). El
tercer carbono del glicerol está unido porintermedio de un grupo
fosfato a una molécula orgánica hidrofílica (afín al agua), que
generalmentecontiene un átomo de nitrógeno o un hidrato de carbono.
Las moléculas de este tipo, con una regiónhidrofóbica y otra
hidrofílica, se denominan moléculas anfipáticas. Todas las
moléculas que conforman elcentro de la membrana tienen
características anfipáticas.
1 Singer, SJ; Nicolson, GL (Febrero de 1972) «The fluid
mosaic model of the structure of cell membranes» Science. Vol. 175.
n.º23. pp. 720 –723.
-
8/17/2019 9 Membranas Biológicas y Transporte
3/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
3 3
Debido a que las moléculas del tipo de los fosfolípidos tienen
un extremo que se asocia libremente con elagua y otro que no lo
hace, cuando se encuentran dispersas en agua adoptan, por lo
general, unaconformación de capa doble. La estructura en bicapa
permite que los grupos del extremo hidrofílico se
asocien libremente con el medio acuoso, y que las cadenas
hidrofóbicas de ácidos grasos permanezcanen el interior de la
estructura, lejos de las moléculas de agua. No todos los lípidos
son capaces de formarbicapas: algunos forman micelas con las colas
hidrocarbonadas hacia el interior, como es el caso de losácidos
grasos libres. Los triglicéridos, por ejemplo, son hidrófobos, de
manera que forman gotas de aceitedentro de la célula. Las
características más importantes de los lípidos que forman bicapas
son, entonces:a) ser claramente dipolares, exhibiendo un polo
hidrofóbico y otro hidrofílico, lo que hace que susmoléculas sean
fuertemente anfipáticas, y b) su forma les permite asociarse con el
agua en forma de unaestructura de doble capa.
Estas bicapas lipídicas tienden a cerrarse sobre si mismas
formando compartimientos herméticos,eliminando así los bordes
libres en los que las colas hidrofóbicas podrían estar en contacto
con el agua.Por esta misma razón los compartimientos formados por
bicapas lipídicas tienden a cerrarse de nuevo
después de haber sido rotos, propiedades denominadas de
autoensamblaje y autosellado. Además unabicapa lipídica tiene
otras características que hacen de ella una estructura ideal para
constituir membranascelulares, de las cuales una de las más
importantes es su fluidez, crucial para muchas funciones.
La bicapa lipídica como un líquido bidimensionalUna
característica importante de las bicapas de fosfolípidos es que en
determinadas condiciones
se comportan como cristales líquidos. Las bicapas tienen
propiedades semejantes a los cristales, perotambién tienen
propiedades semejantes a los líquidos, porque a pesar de la
ordenada disposición de susmoléculas los grupos hidrocarbonados
están en movimiento constante. Por lo tanto, una molécula
sedesplaza rápidamente de un punto al otro en un mismo lado de la
estructura. Este movimiento confiere a labicapa la propiedad de un
fluido bidimensional '. En condiciones normales, esto
significa que una molécula
de fosfolípido puede atravesar la superficie de una célula
eucariota en pocos segundos.Las propiedades de líquido de la bicapa
lipídica también permiten el desplazamiento de las
moléculas que se encuentran insertas en ella sobre el planode la
membrana (siempre que no estén ancladas mediantealgún otro
mecanismo, como en algunas conexionesintercelulares).
La fluidez de las membranas celulares esbiológicamente
importante. Algunos procesos de transporte yactividades enzimáticas
pueden detenerse cuando laviscosidad de la membrana (parámetro
inversamenterelacionado con la fluidez) se incrementa más allá de
un nivelcrítico umbral. La fluidez de la bicapa depende tanto de su
composición como de la temperatura. Unamenor longitud de las
cadenas reduce la tendencia de las colas a interaccionar entre sí y
los doblesenlaces cis producen pliegues en las cadenas
hidrocarbonadas que dificultan su empaquetamiento, deforma que las
membranas permanecen fluidas a temperaturas más bajas.
La mayor parte de las membranas biológicas se encuentran en un
estado cristalino líquido; sinembargo, a temperaturas bajas, las
fuerzas de Van der Waals1 entre las cadenas de
hidrocarburosdispuestas una cerca de otra convierten las bicapas de
fosfolípidos en un gel sólido. Bacterias, levaduras yotros
organismos cuyas temperaturas varían con la de su entorno controlan
la composición de ácidosgrasos de sus lípidos de membrana para
mantener una fluidez relativamente constante. Si la temperatura
1 Ocurren entre dipolos inducidos no permanentes de
enlaces covalentes poco polares, como C H. Cuando los
orbitalesmoleculares de dos enlaces apolares se ponen en contacto,
las nubes de electrones se rechazan mutuamente y se inducendipolos,
lo que establece una fuerza de atracción débil. Conforme los átomos
se alejan, los dipolos desaparecen y se pierde lafuerza de van der
Waals
1 Singer, SJ; Nicolson, GL (Febrero de 1972) «The fluid
mosaic model of thestructure of cell membranes» Science. Vol. 175.
n.º 23. pp. 720 –723.
-
8/17/2019 9 Membranas Biológicas y Transporte
4/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
4 4
disminuye sintetizan ácidos grasos insaturados, de manera de
evitar la pérdida de fluidez de susmembranas por efecto de la
disminución de la temperatura.
Los principales fosfolípidos que se encuentran en las membranas
son la fosfatid i lcolina,
fosfatidiletanolamina, fosfatidilserina y esfingomielina.
La razón por la cual hay tal variedad de fosfolípidosquizás se deba
a que serían necesarios para los distintos tipos de proteínas que
se hallan asociadas a lamembrana, que únicamente podrían funcionar
en presencia de grupos polares específicos, como los quebrinda la
cabeza polar de los distintos tipos de fosfolípidos. Algunos
fosfolípidos como el fosfatidilinositol,son funcionalmente
importantes pero se hallan en cantidades relativamente pequeñas A
menudo lasmembranas plasmáticas bacterianas están compuestas por un
único tipo de fosfolípido y no contienencolesterol. Contrariamente,
la composición de la membrana celular de la mayoría de las
célulaseucarióticas es más variada conteniendo además algún
esterol (colesterol en las células animales)
y glicolípidos. Las membranas plasmáticas de algunas
células animales contienen cantidadesespecialmente elevadas de
colesterol, hasta una proporción de más de una molécula de
colesterol porcada molécula de fosfolípido. Las moléculas de
colesterol se acomodan entre los fosfolípidos y actúan
como "amortiguadores de fluidez". A bajas temperaturas las
moléculas de colesterol se interponen entrelas cadenas de
hidrocarburos, con lo cual evitan quese acerquen y formen
interacciones de van derWaals, los cuales provocarían la
cristalización de lamembrana. Por otro lado, a temperaturas
elevadas,las moléculas de colesterol restringen el
movimientoexcesivo de las cadenas de ácidos grasosdisminuyendo la
fluidez de la membrana.
Las bicapas lipídicas, sobre todo las que seencuentran en estado
de cristal líquido, tienentambién otras propiedades biológicas
importantes.
Las bicapas tienden a resistir la formación deextremos libres:
como resultado tienden aautosellarse y casi en cualquier
circunstancia formanvesículas cerradas espontáneamente. Por último,
en
condiciones apropiadas, las bicapas son capaces de fusionarse
con otras. La fusión de membranas es unfenómeno celular muy
importante y que requiere de membranas en estado fluido para
producirse. Cuandouna vesícula se fusiona con otra membrana, ambas
bicapas y sus compartimientos forman unacontinuidad. Esto permite
tanto la transferencia de material de un compartimiento a otro, o
el movimientode una vesícula secretora hacia afuera de la célula,
mediante un proceso Ilamado exocitosis. De modosimilar, aunque
inverso, la endocitosis permite la incorporación de grandes
moléculas del exterior mediantela formación de vesículas en alguna
porción de la membrana.
Otro aspecto importante es que la bicapa lipídica es asimétrica,
hecho que tiene una obviarelación funcional y se refiere a la
diferente composición lipídica de cada una de sus monocapas. En
losglóbulos rojos, la mayoría de las moléculas lipídicas que tienen
colina en su grupo cabeza (fosfatidilcolinay esfingomielina) se
encuentran en la mitad exterior de la bicapa, mientras que la
mayoría de lasmoléculas de fosfolípido que contienen un grupo amino
(fosfatidiletanolamina y fosfatidilserina) se hallanen la mitad
interior. Algunos fosfolípidos, como el fosfatidilinositol, actúan
como intermediarios en elproceso de señalización celular: ante
estímulos extracelulares, el fosfatidilinositol ubicado en el
interior dela membrana es primero fosforilado en dos oportunidades
y luego hidrolizado en trifosfato de inositol ydiacilglicerol.
Ambos fragmentos de la molécula actúan dentro de la célula como
mensajeros solubles quepermiten la difusión de la señal hacia el
interior de la célula.
Como ya se ha mencionado, la síntesis de fosfolípidos ocurre en
el retículo endoplásmico liso y esallí donde se genera la asimetría
por traslocadores que trasladan específicamente moléculas
defosfolípidos de una capa a la otra. Los fosfolípidos son
sintetizados en la cara externa (citosólica). Si bien
-
8/17/2019 9 Membranas Biológicas y Transporte
5/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
5 5
se sintetizan tanto fosfatidilcolina como el resto de los
fosfolípidos, sólo la fosfatidilcolina pasa a la carainterna de la
membrana del retícu lo gracias a un traslocador fosfolipídico
específico (una “flipasa”). La
pérdida de la asimetría de la membrana es una señal de
muerte celular, ya que la exposición de
fosfatidilserina en la monocapa externa es un índice de
apoptosis o muerte celular programada quefavorece la fagocitosis de
estas células por macrófagos.
También existen glicolípidos en la membrana. Se presentan
probablemente en las membranasplasmáticas de todas las células
animales, constituyendo el 5% de las moléculas de lípido de la
monocapaexterna y son las moléculas que presentan una asimetría mas
marcada en cuanto a su distribución en lasmembranas celulares.
Estas moléculas se encuentran exclusivamente en la mitad no
citoplasmática de lamembrana plasmática. En la membrana plasmática,
los grupos azúcares quedan al descubierto en lasuperficie de la
célula, lo que sugiere que deben desempeñar una función en las
interacciones de la célulacon su entorno. Debido a que los azúcares
se añaden en la cara luminal del aparato de Golgi, al formarsela
vesícula de transporte el residuo glicosídico queda hacia el
interior de la misma, pero cuando ésta sefusiona con la membrana
plasmática, la porción glicosilada, que es hidrófila, queda hacia
el exterior de la
célula. Hay variados tipos de glicolípidos: los más complejos
contienen oligosacáridos con uno o másresiduos de ácido siálico que
les proporciona carga negativa. Estos lípidos son más abundantes en
lamembrana plasmática de células nerviosas.
La función de los glicolípidos puede ser variada: en las células
epiteliales tapizan la cara que da alepitelio, donde las
condiciones son extremas (bajos o altos valores de pH, enzimas
degradativas),protegerían la integridad de las propias proteínas de
membrana; también cumplen funciones aislantes,como ocurre en la
membrana que rodea el axón de las células nerviosas, tapizada
totalmente porglicolípidos en la cara externa. La presencia de
carga eléctrica negativa en su molécula es responsabletambién de la
concentración de iones, especialmente Ca+2 en la superficie
externa. Además desempeñanuna importante función en procesos de
reconocimiento celular, ayudando a su vez a las células a unirse
ala matriz extracelular y a otras células.
Proteínas de membranaAunque la estructura básica de las
membranas biológicas es provista por los fosfolípidos, la
mayoría de las funciones específicas de la membrana son llevadas
a cabo por proteínas. De acuerdo conello, las cantidades y tipos de
proteínas de membrana son muy variables: en la membrana mielínica,
quesirve de aislación eléctrica al axón de la neurona, menos del
25% son proteínas, en tanto que en lasmembranas donde hay
transducción energética (mitocondrias y cloroplastos) el porcentaje
alcanza al 75%.En promedio, hay un 50% de lípidos y otro tanto de
proteínas, pero como las proteínas son mucho másgrandes, la
relación numérica es de alrededor de 50 moléculas de
fosfolípidos por cada molécula deproteína.
Al principio, los investigadores encontraban difícil pensar que
las proteínas se asociaran en otrositio que no fuese la superficie
exterior de las membranas. Sin embargo en estudios fisicoquímicos
de lasproteínas de membrana se mostró que gran parte de éstas son
del tipo globular; esto significa que sondemasiado voluminosas como
para asociarse sólo a la superficie de las membranas. Por último,
seobtuvieron pruebas por las cuales puede afirmarse que algunas
proteínas se asocian con las membranasde tal manera que una región
(o dominio) de ellas se encuentra de un lado de la membrana y otra
en ellado opuesto. Por lo tanto, el modelo de la estructura de
membrana más razonable es aquél en el que seforma un mosaico de
proteínas, en el cual gran parte de éstas son móviles y se
extienden dentro o aambos lados de la bicapa lipídica.
Hoy se sabe que existen dos tipos de proteínas de
membrana: proteínas integrales ointrínsecas y proteínas
periféricas o extrínsecas. Las proteínas integrales de membrana
poseen
algunas regiones insertadas en las regiones hidrófobas de la
bicapa lipídica. Algunas atraviesan toda lamembrana, de manera que
gran parte de ellas se encuentra en alguno de los lados de la
membrana; estasproteínas integrales se llaman también proteínas
transmembrana. Algunas otras proteínas integrales
-
8/17/2019 9 Membranas Biológicas y Transporte
6/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
6 6
poseen solo una pequeña porción dentro de la bicapa que sirve
como ancla y el resto de la molécula en elcitoplasma o hacia la
superficie celular. Otras están casi por completo insertas en la
región hidrófoba yposeen cadenas polipeptídicas que atraviesan una
y otra vez (incluso hasta doce veces) la bicapa lipídica.
Las proteínas integrales de membrana son capaces de insertarse
en la bicapa lipídica debido aque las porciones que lo hacen son
hidrófobas y, por tanto, son compatibles con el interior de
lamembrana. Cuando alguna proteína de membrana contiene una porción
hidrofílica, ésta generalmente seencuentra en protrusión por fuera
de la superficie de la membrana, en contacto con el medio acuoso
2.
También las proteínas transmembránicas pueden estar unidas por
una cadena de ácido graso a lacara interna de la membrana, pasar
una sola vez a través de la misma (1) o muchas veces (2),
estarsolubles en el citosol pero ancladas a la cara interna por un
resto acilo3 o prenilo (3), o haber sidosintetizadas como
proteínas transmembrana en el retículo endoplásmico rugoso y unidas
a un resto deglicosilfosfatidilinositol, que las ancla a la cara
externa de la membrana (4).
En las proteínas integrales, la parte que se halla dentro de la
membrana usualmente adopta unaestructura en -hélice, con predominio
de aminoácidos hidrofóbicos. Además este enrollamiento aseguraque
todas las uniones peptídicas (que son polares) estén disminuidas en
su polaridad debido a laformación de puentes de hidrógeno. Sólo
hacen falta 20-30 aminoácidos para atravesar la membrana enforma de
-hélice y sólo unos diez para hacerlo en forma de hoja -plegada. La
mayoría de las proteínasintegrales atraviesan la membrana en forma
de hélices , pero algunas (como las porinas de bacterias yde
mitocondrias) forman láminas que se disponen en forma de barril (
“-bar rel”). Las proteínasintegrales de membrana pueden ser
solubilizadas por medio de detergentes, que en agua forman
micelas.
El otro tipo de proteínas de membrana, las proteínas
periféricas, pueden eliminarse de ésta sinalterar la estructura de
la doble capa. Por lo general se unen a regiones expuestas de
proteínas integrales.Las proteínas periféricas están usualmente
asociadas por interacciones no covalentes a otras
proteínastransmembránicas y se pueden ubicar hacia adentro (5) o
hacia fuera (6) de la membrana.
Si bien las proteínas pueden migrar dentro de la membrana,
existen distintos dispositivos por loscuales las células pueden
confinar a las proteínas dentro de determinados dominios
membranosos. En lascélulas epiteliales que tapizan el intestino y
los túbulos renales, las proteínas que miran hacia la luz del
2 La diferencia entre proteínas solubles y proteínas
ligadas a membranas no consiste en que una contenga
aminoácidoshidrófobos y la otra no, sino en que en aquéllas los
aminoácidos hidrófobos se encuentran en el interior de la molécula,
lejos
del medio acuoso, en tanto que en éstas las porciones hidrófobas
de los aminoácidos se presentan en la superficie de lamolécula, en
contacto con las cadenas de ácidos grasos de la bicapa lipídica
3 Un grupo acilo es un grupo derivado de un oxoácido,
normalmente un ácido carboxílico, por eliminación de al menos un
grupohidroxilo. Los derivados de un ácido carboxílico tienen como
fórmula general R-CO-.
-
8/17/2019 9 Membranas Biológicas y Transporte
7/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
7 7
tubo no pueden pasar hacia las paredes laterales o hacia la cara
opuesta de la célula debido a laexistencia de uniones estrechas (se
verán más adelante) que les impiden el paso. Sin embargo no es
elúnico medio, aunque en algunos casos no se conoce el mecanismo
que impide la migración libre: en
espermatozoides de mamíferos las proteínas del ápice, del resto
de la cabeza y de la cola forman tresdominios perfectamente
individualizables con anticuerpos fluorescentes, pero se desconoce
cuál es lafunción de dichas proteínas y las fuerzas que impiden su
migración. Otros ejemplos de confinamiento deproteínas en
determinadas zonas de la membrana están dadas por asociación de
proteínas con elcitoesqueleto (glóbulos rojos), o con la matriz
extracelular, o con ambos, o puede haber interacciones
entreproteínas de dos células distintas en zonas de membrana
contiguas.
Asimetría de proteínas de las membranasUna de las pruebas más
evidentes de que las proteínas se insertan en la bicapa lipídica
proviene
del estudio con microscopía electrónica por el método de
criofractura, el cual prácticamente permite a losinvestigadores
observar las membranas de "adentro hacia afuera". Al comparar, con
el método descrito,
las dos superficies de una membrana, se muestra que en una de
estas superficies hay gran cantidad departículas, en tanto que en
la otra superficie se observan muy pocas. Estas partículas son
proteínasembebidas en la doble capa de lípidos. De aquí se deduce
que las proteínas presentes en una membranabiológica están
distribuidas de manera asimétrica. Cada lado de la membrana tiene
característicasdiferentes debido a que cada proteína de membrana se
orienta en la bicapa en un solo sentido. Estaasimetría está dada
por la forma tan específica en que se forman e intercambian las
membranas de unaparte de la célula a otra.
Los hidratos de carbono se encuentran unidos a las porciones de
las proteínas expuestas en lasuperficie celular, y no a las que se
internan al citoplasma. Esta distribución asimétrica de los
hidratos decarbono se debe a la forma en que las glucoproteínas se
insertan en las membranas al ser sintetizadas.Como se dijo, las
proteínas de la membrana plasmática son producidas por ribosomas
del retículo
endoplásmico rugoso (RER) y se insertan en la membrana de éste
durante su síntesis. Los hidratos decarbono se añaden a las
proteínas en el lumen del RER. Si se sigue de cerca la gemación y
fusión demembrana que forman parte del proceso de transporte se
podrá observar que la porción proteica enprotrusión hacia el
compartimiento interno (cisterna) del RER también estará expuesta
al interior delcomplejo de Golgi, donde se encuentran las enzimas
que modifican los hidratos de carbono de lasproteínas. La porción
proteica permanecerá en el compartimiento interior al separarse del
complejo deGolgi para ser empaquetada en una vesícula secretora.
Cuando la vesícula secretora se fusiona con lamembrana plasmática,
la porción de la proteína que contiene el hidrato de carbono,
orientado hacia elinterior de la vesícula, se convertirá en parte
de la proteína de membrana expuesta en la superficie celular.
Funciones de las proteínas de membrana ¿Por qué motivo la
membrana plasmática requiere de tantas proteínas distintas? La
diversidad de
proteínas en una membrana refleja el número de funciones que se
llevan a cabo en ella.Habitualmente la manera en que una proteína
se asocia a la bicapa lipídica es un indicativo de la
función de la proteína. Así, sólo las proteínas transmembrana
pueden actuar a ambos lados de la bicapa otransportar moléculas a
través de ellas. Las proteínas de la membrana plasmática pueden ser
clasificadasen diferentes grupos, de acuerdo a la función que
desempeñan:a) Proteínas de adhesión celular, que unen
firmemente las membranas de células adyacentes y actúan
como puntos de anclaje con componentes del
citoesqueleto.b) canales proteicos entre dos células (uniones
de hendidura, permiten el paso de moléculas pequeñas
entre dos células vecinas.
c) Proteínas de transporte que permiten el transporte
selectivo de moléculas esenciales, ya sea enforma pasiva o en forma
activa mediante procesos que requieren de energía.
-
8/17/2019 9 Membranas Biológicas y Transporte
8/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
8 8
d) Proteínas transductoras receptoras de señales, que se
unen a moléculas portadoras de señalesexternas y que luego
transfieren el mensaje al interior de la célula.
e) Bombas dependientes de ATP, que transportan activamente
iones de un compartimiento a otro,
constituyendo así un mecanismo de almacenamiento de energía (se
verán en mitocondrias ycloroplastos).f) Algunas proteínas
intrínsecas de la membrana actúan como enzimas, con sitios activos
localizados
en la superficie de la membrana o en el interior de ella.
PASO DE LOS MATERIALES A TRAVÉS DE LAS MEMBRANAS (Transporte de
membrana)Debido a su interior hidrofóbico, la bicapa lipídica de
una célula constituye una barrera altamenteimpermeable a la mayoría
de las moléculas polares. Esta función de barrera es de especial
importancia, yaque permite a una célula mantener en su citosol
ciertos solutos a concentraciones diferentes a las queestán en el
fluido extracelular y en cada uno de los compartimientos
intracelulares. Sin embargo, parapoder utilizar esta barrera las
células han tenido que desarrollar sistemas para transportar
específicamentemoléculas hidrosolubles a través de la membrana y
así poder ingerir nutrientes esenciales, excretar losproductos de
deshecho del metabolismo y regular las concentraciones
intracelulares de iones. El
transporte de iones inorgánicos y de pequeñas moléculasorgánicas
hidrosolubles a través de la bicapa lipídica seconsigue mediante
proteínas transmembrana especializadas,
cada una de las cuales es responsable de la transferencia deuna
molécula o un ion específico o de un grupo de moléculasafines.
El hecho de que una membrana permita el paso deciertas
sustancias depende de su estructura y del tamaño ycarga eléctrica
de las moléculas. Se dice que una membrana espermeable para
alguna sustancia si permite que ésta la cruce eimpermeable si
no permite el paso de dicha sustancia. Unamembrana selectivamente
permeable permite el paso dealgunas sustancias pero no el de
otras. Todas las membranasbiológicas (aquellas que rodean a las
células, núcleos,
vacuolas, mitocondrias, cloroplastos y otros organeloscelulares)
son selectivamente permeables.
-
8/17/2019 9 Membranas Biológicas y Transporte
9/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
9 9
Al reaccionar a las condiciones ambientales cambiantes o a las
diversas necesidades de la célula,la membrana puede constituir una
barrera al paso de un compuesto determinado en cierto
momento,mientras promueve activamente su paso en otro momento.
Mediante la regulación del tráfico químico de
esa manera, la célula controla su propia composición interna de
iones y moléculas que puede ser muydiferente a la del exterior. En
el mundo abiótico, los materiales se mueven pasivamente por
procesosfísicos como la difusión. En los seres bióticos, los
materiales también se mueven activamente por procesosfisiológicos
como transporte activo, exocitosis y endocitosis. Esos
procesos fisiológicos activosdemandan un gasto de energía por parte
de la célula.
DifusiónAlgunas sustancias se desplazan hacia adentro y afuera
de las células, y se mueven dentro de
éstas por medio de un proceso llamado difusión simple, el cual
se basa en el desplazamiento al azar.Cuando se deja caer un terrón
de azúcar en un vaso de precipitado lleno con agua, las moléculas
de aquélse disuelven y luego comienzan a difundirse hacia toda el
agua del recipiente, como consecuencia de que
las moléculas de azúcar individuales se desplazan al azar en
todas direcciones. En última instancia, ladifusión ocasiona una
distribución uniforme de las moléculas de azúcar en toda el agua
del vaso. Por lotanto, puede decirse que la difusión implica el
movimiento neto de partículas en favor de un gradiente
deconcentración (diferencia de concentración de una sustancia de un
punto a otro).
La velocidad de difusión está en función del tamaño y forma de
las moléculas, de sus cargaseléctricas y de la temperatura. Al
aumentar la temperatura, las moléculas se mueven con mayor rapidez
yaumenta la proporción de difusión. Este movimiento de las
partículas pequeñas (llamado movimientobrowniano) constituye un
modelo que explica el mecanismo de difusión de las moléculas.
DiálisisLa difusión de un soluto (una sustancia disuelta) a
través de una membrana diferencialmente
permeable se llama diálisis. Para demostrar la diálisis se
utiliza una bolsa de celofán, llena con unasolución de azúcar, que
luego se sumerge en un matraz que contiene agua pura. Si la
membrana decelofán es permeable a azúcar y al agua, las moléculas
de azúcar pasarán a través de ella hasta que laconcentración de
azúcar en el agua de los dos lados de la membrana sea exactamente
igual. A partir deese momento, las moléculas de soluto (así como
también las moléculas de agua) seguirán pasando através de la
membrana, pero ya no habrá ningún cambio neto en las
concentraciones, ya que la velocidadde movimiento será igual en
ambos sentidos. La diálisis renal es una aplicación práctica de
este proceso;los productos de desecho, que se difunden a través de
las membranas artificiales del aparato, puedenretirarse del
organismo, pero los eritrocitos, proteínas sanguíneas y otras
moléculas grandes, no sedifunden a través de la membrana y por
tanto se retendrán en el organismo.
El celofán con frecuencia se utiliza como "membrana artificial"
Está compuesto por moléculas depolisacáridos y puede formar una
lámina delgada que permite el paso de moléculas de agua.
Estasmembranas se fabrican con permeabilidad variable para
diferentes solutos.
Con suficiente tiempo, prácticamente cualquier molécula acabará
difundiendo a través de la bicapalipídica. Sin embargo la velocidad
a la que se produce esta difusión varía enormemente dependiendo
enparte del tamaño de la molécula y principalmente de su
solubilidad relativa. En general cuanto menor ymenos soluble en
agua sea una molécula (es decir, cuanto más hidrofóbica o no polar)
más rápidamentedifunde a través de la bicapa. Las moléculas
pequeñas no polares (O2, CO2) se disuelven fácilmente en labicapa
lipídica y por lo tanto difunden con rapidez. Las moléculas polares
no cargadas también difundenrápidamente a través de la bicapa
lipídica si su tamaño es reducido: por ejemplo agua, etanol y
ureaatraviesan rápidamente una bicapa, el glicerol lo hace con
menor rapidez y la glucosa prácticamente no la
atraviesa.Las moléculas de agua, por ejemplo, pueden desplazarse
fácilmente a través de una bicapalipídica fluida, pasando a través
de brechas que se forman cuando una cadena de ácido graso se
mueve
-
8/17/2019 9 Membranas Biológicas y Transporte
10/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
10 10
momentáneamente. Por el contrario, las bicapas lipídicas son
altamente impermeables a todas lasmoléculas cargadas (iones) por
muy pequeñas que sean; la carga y el grado de hidratación les
impidepenetrar la fase hidrocarbonada de la bicapa.
ÓsmosisLa ósmosis es una variedad especial de difusión y en
consecuencia transporte pasivo, que implica
el movimiento de moléculas solventes (en este caso, el agua) a
través de una membrana de permeabilidadselectiva. Las moléculas de
agua pasan libremente en cualquier dirección, pero al igual que en
todos losprocesos de difusión, el movimiento neto ocurre a partir
de la región de mayor concentración a la demenor, separadas por una
membrana semipermeable, para igualar concentraciones. De acuerdo al
medioen que se encuentre una célula, la ósmosis varía. Dicho
proceso no requiere gasto de energía.
Permeabilidad de la bicapa lipídica a diferentes sustancias
Tipo de molécula Ejemplo PermeabilidadHidrófoba
N2, O2• hidrocarburos Permeabilidad librePolar pequeña
H20, CO2, glicerol, urea Permeabilidad librePolar grande
Glucosa y otros monosacáridos
y disacáridos sin cargaNo permeable
Iones y moléculas con carga Aminoácidos, H+, HCO-3, Ca+,Cl-;
Mg+
No permeable
Los principios que intervienen en el proceso de ósmosis se
ilustran mediante la utilización de unaparato llamado tubo en U. El
tubo en U se divide en dos secciones por una membrana de
permeabilidad
selectiva que impide el paso de las moléculas de soluto
(glucosa, sal y otras) En una parte del tubo secoloca una solución
de agua y solutos; en la otra se coloca agua pura. La solución de
agua y solutoscontiene una concentración de agua menor a la del
agua pura, porque las moléculas de soluto han"diluido" las
moléculas de agua. Por tanto, hay un movimiento neto de moléculas
de agua del lado delagua pura (con mayor concentración de moléculas
de agua) hacia el lado del agua con soluto (que tienemenor
concentración de moléculas de agua), como resultado de esto el
nivel de líquido del lado del aguapura disminuye, mientras que se
eleva el del lado del agua con soluto. Sin embargo, aún existe
unadiferencia en la concentración de las moléculas de agua entre
ambos lados, debido a que las moléculas desoluto no pueden moverse
a través de la membrana. El movimiento neto de agua continuará, y
el nivel delíquido del lado del agua con soluto seguirá aumentando.
En condiciones no sujetas a la gravedad, esteproceso continuaría
indefinidamente, pero en la tierra, el peso de la columna de
líquido en aumento
finalmente ejercerá una presión suficiente para detener el
cambio en los niveles de líquido, aunque lasmoléculas de agua
continuarán pasando a través de la membrana en ambas direcciones.La
presión osmótica de una solución está relacionada con la tendencia
que presenta el agua de
moverse hacia dicha solución mediante ósmosis. En el ejemplo del
tubo en U se podría medir la presiónosmótica insertando un pistón
del lado del agua con soluto y midiendo la presión necesaria que
debeejercer el pistón para evitar el aumento en el nivel del
líquido de dicho lado del tubo. Una solución con unaalta
concentración de soluto tendrá una baja concentración de agua y una
elevada presión osmótica; encambio, una solución con una baja
concentración de soluto tendrá una elevada concentración de agua
yuna baja presión osmótica.
Soluciones isotónicas, hipertónicas e hipotónicas
Con frecuencia deseamos comparar las presiones osmóticas de dos
soluciones. En todocompartimiento de una célula viva se encuentran
disueltas sales, azúcares y otras sustancias que leconfieren a
dicho líquido una determinada presión osmótica. Cuando una célula
se coloca en una solución
-
8/17/2019 9 Membranas Biológicas y Transporte
11/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
11 11
cuya presión osmótica es igual a la suya, no hay movimiento neto
de moléculas de agua, ni hacia afuera nihacia adentro de ella; por
tanto, la célula no se hincha ni se encoge. Se dice que el líquido
en el cual secolocó la célula es un líquido isotónico (es
decir que tiene presión osmótica igual) con respecto al líquido
del interior de la célula. Normalmente, el plasma de la sangre
(componente líquido de ella) y los demáslíquidos corporales son
isotónicos con respecto al líquido intracelular; es decir,
contienen unaconcentración de agua igual a la del líquido
intracelular. Una solución de cloruro de sodio al 0,9%
(llamadasolución salina fisiológica) es isotónica respecto a las
células humanas y a las células de otros mamíferos.Los eritrocitos
humanos colocados en una solución de cloruro de sodio al 0,9% no se
encogen ni sehinchan.
Concentración de solutosen la Solución A
Concentración desolutos en la Solución B
Tonicidad Dirección demovimiento del agua
Mayor Menor A es hipertónica respecto a B;B es hipotónica
respecto a A
B hacia A
Mayor Mayor B es hipertónica respecto a A.
A es hipotónica respecto a B
A hacia B
Igual Igual Isotónica No hay movimientoneto
Si el líquido circundante tiene unaconcentración de solutos
mayor que la dellíquido intracelular y, por tanto, una
presiónosmótica mayor que la de éste se dice que esuna solución
hipertónica; una célula colocadaen una solución hipertónica pierde
agua y portanto, se encoge. Así, los eritrocitos humanos
colocados en una solución de cloruro de sodioal 1,3% pierden
agua y se encogen. Cuandouna célula con pared celular se coloca en
unmedio hipertónico pierde agua, y entonces sucontenido disminuye
dentro de la pared celular;este proceso se llama plasmólisis.
Dichofenómeno se observa en las plantas cuando sedepositan grandes
cantidades de sales ofertilizantes en la tierra o agua que las
rodea. Si
el líquido circundante posee una concentración de solutos menor
que la del líquido intracelular, y por lotanto tiene una presión
osmótica menor que la de éste, se denomina solución hipotónica; en
estascircunstancias, el líquido se desplazará hacia el interior de
la célula, provocando que ésta se hinche,ejerciendo presión contra
las membranas celulares llamada presión de turgencia, las cuales se
ponentensas, fenómeno llamado turgencia (del latín turgens-
turgentis; hinchar). En una solución de cloruro desodio al 0,6%, el
agua entra hacia el interior de los eritrocitos, provocando el
hinchamiento de éstos yeventualmente su ruptura o lisis
(visualizado por la disolución de la hemoglobina en el líquido
hipotónico,que lo colorea de rojo). Las paredes celulares rígidas
de células vegetales, algas, bacterias y hongoshacen posible que
esos organismos vivan sin reventar en un medio externo muy diluido,
que contenga unaconcentración muy baja de solutos. Debido a las
sustancias disueltas en el citoplasma, las células sonhipertónicas
respecto al medio externo (el medio circundante es hipotónico
respecto al citoplasma.). Elagua tiende a difundir hacia el
interior de las células por ósmosis, llenando sus vacuolas
centrales ydistendiéndolas. La presión de turgencia es un
factor importante en el sostén del cuerpo de las plantasherbáceas.
Por este motivo, una flor se marchita cuando la presión de
turgencia de sus células disminuye(las células han sufrido
plasmólisis) por falta de agua.
EN SOLUCI N
ISOTÓNICA
EN SOLUCI N
HIPERTÓNICA
EN SOLUCI N
HIPOTÓNICA
-
8/17/2019 9 Membranas Biológicas y Transporte
12/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
12 12
Transporte mediado de moléculas pequeñasLa membrana celular es
relativamente impermeable a casi todas las grandes moléculas
polares.
Esto constituye una ventaja biológica para la célula, ya que
casi todos los compuestos metabolizados ensu interior son polares y
la impermeabilidad de la membrana impide su pérdida por difusión.
Paratransportar nutrientes polares, como glucosa y aminoácidos, a
través de la membrana lipídica hacia elinterior de la célula, han
aparecido a lo largo de la evolución sistemas de proteínas
transportadoras que seunen a esas moléculas y facilitan su pasaje a
través de la membrana. El paso de solutos a través de lamembrana
celular por el sistema de transporte se llama transporte
mediado.
Transporte Pasivo (Difusión facilitada)Algunas moléculas son
demasiado grandes como para difundir a través de los canales de
la
membrana y demasiado insolubles en lípidos como para poder
difundir a través de la capa de fosfolípidos.Tal es el caso de
azúcares como la glucosa, aminoácidos, nucleótidos y muchos
metabolitos. En los casos
más simples, la célula utiliza la energía almacenada por el
gradiente de concentración de una sustancia
cuya concentración es mayor en el líquido extracelular que en el
intracelular. En estas circunstancias,mientras la membrana sea
permeable a dicha sustancia, ésta se desplazará hacia el interior
de la célula.Para que esto suceda se necesita de la presencia de
proteínas especiales de membrana. Este tipo detransporte se llama
transporte pasivo o difusión facilitada.
La difusión facilitada depende de la existencia de
proteínas transportadoras, las cuales secombinan temporalmente con
la molécula de soluto para acelerar el paso de ésta a través de la
membranacelular. Cada proteína está destinada al transporte de un
tipo particular de molécula y con frecuencia deuna cierta especie
molecular, por lo que presentan especificidad. La proteína
transportadora no se
modifica por esta acción; después de transportar la molécula de
soluto, queda libre para unirse a unanueva molécula. Todas las
proteínas de transporte estudiadas son proteínas transmembranales.
Ladifusión facilitada es mucho más rápida que la difusión simple y
depende:
Comparación entre un transporte pasivo a favor del gradiente
electroquímico y el transporteactivo en contra del gradiente
electroquímico
-
8/17/2019 9 Membranas Biológicas y Transporte
13/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
13 13
* Del gradiente de concentración de la sustancia a ambos lados
de la membrana* Del número de proteínas transportadoras existentes
en la membrana* De la rapidez con que estas proteínas hacen su
trabajo
El transporte de las moléculas deglucosa en los eritrocitos es
un buenejemplo de difusión facilitada portransportador. Las
moléculas quetransportan glucosa son glucoproteínas;éstas
comprenden un 2% de las proteínastotales de la membrana. Las
célulasmantienen una baja concentración internade glucosa mediante
la adición inmediatade un fosfato a las moléculas de glucosaque
entran en ellas: de esta manera
convierten dichas moléculas en fosfatosde glucosa con elevada
carga eléctrica y,así, no pueden regresar al otro lado de
lamembrana. El mecanismo de transportede glucosa no se comprende a
fondo.Parece que la proteína transportadora noforma un “hoyo” en la
membrana para quela glucosa pase a través de él; si tal fuerael
caso, otras moléculas similares a la glucosa y algunas moléculas
más pequeñas que ésta tambiénpodrían pasar a través del "hoyo". Al
parecer, lo que ocurre es que la glucosa se une de modo específico
auna porción de proteína expuesta en la superficie celular externa,
y con esto, modifica la conformación de
la proteína, de manera que se abre un canal dentro de la
proteína misma (o entre varias subunidades de lamisma cadena
polipeptídica), que permite el paso de la molécula de glucosa para
liberarla en el interior dela célula. Según este modelo, una vez
que la glucosa se libera en el interior de la célula, la
proteínarecupera su conformación original y está lista para unirse
nuevamente a una molécula de glucosa en lasuperficie celular. Otra
clase de proteínas transportadoras son las formadoras de canal;
estas no se unenal soluto sino que forman poros hidrofílicos
que atraviesan la bicapa lipídica, que al estar abiertospermiten
que determinados solutos (habitualmente iones inorgánicos de tamaño
y carga apropiados)puedan pasar a su través y por lo tanto
atravesar la membrana).
Transporte activoAlgunas moléculas se transportan a través de la
célula mediante el proceso de difusión; a otras las
requiere la célula en concentraciones mayores a su concentración
extracelular. Estas moléculas seincorporan mediante mecanismos
detransporte activo. Este mecanismoexige una fuente de energía
debido aque el transporte activo implica el"bombeo" de una molécula
en contra desu gradiente de concentración (de unazona de baja
concentración hacia unade concentración elevada). Por tanto,los
sistemas de transporte activo
utilizan energía generada por elmetabolismo celular en forma
detrifosfato de adenosina (ATP) o bien
-
8/17/2019 9 Membranas Biológicas y Transporte
14/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
14 14
utilizan algún otro tipo de energía almacenada, derivada de la
hidrólisis del ATP. Los sistemas detransporte activo están basados
en permeasas específicas e inducibles. Las permeasas son
enzimas quetransportan sustancias a través de la membrana celular,
sea hacia el interior o hacia el exterior de la célula
al mismo tiempo que rompen por hidrolisis un enlace fosfato del
ATP.Un ejemplo de transporte activo es la bomba de calcio esta
es una proteína de la membranacelular de todas las células
eucariotas. Su función consiste en transportar calcio iónico (Ca
2+) hacia elexterior de la célula, gracias a la energía
proporcionada por la hidrólisis de ATP, con la finalidad demantener
la baja concentración de Ca2+ en el citoplasma que es unas
diez mil veces menor que en elmedio externo, necesaria para el
normal funcionamiento celular. Se sabe que las variaciones en
laconcentración intracelular del Ca2+ (segundo mensajero) se
producen como respuesta a diversosestímulos y están involucradas en
procesos como la contracción muscular, la expresión genética,
ladiferenciación celular, la secreción, y varias funciones de las
neuronas. Dada la variedad de procesosmetabólicos regulados por el
Ca2+, un aumento de la concentración de Ca2+ en el citoplasma
puedeprovocar un funcionamiento anormal de los mismos. Si el
aumento de la concentración de Ca2+ en la fase
acuosa del citoplasma se aproxima a un décimo de la del medio
externo, el trastorno metabólico producidoconduce a la muerte
celular. El calcio es el mineral más abundante del organismo,
además de cumplirmúltiples funciones.
Uno de los ejemplos más sorprendentes de los mecanismos de
transporte activo es la bomba desodio y potasio que se
observa en todas las células animales. Esta bomba consta de una
proteínaespecífica, localizada en la membrana plasmática, que
utiliza ATP para intercambiar iones de sodio delinterior de la
célula por iones de potasio de su exterior. Esto provoca un
desequilibrio en la concentraciónde iones de sodio y potasio en los
lados opuestos de la membrana, de manera que en
condicionesnormales, la concentración de potasio sea de 10 a 15
veces mayor en el interior que en el exterior de lacélula y a la
inversa para el sodio. Las células son capaces de utilizar estos
enormes gradientes deconcentración para generar un potencial
eléctrico (separación de cargas eléctricas) a través de la
membrana, el cual constituye la base para la transmisión de los
impulsos eléctricos necesarios para latransmisión de los impulsos
nerviosos. Estos gradientes de concentración también almacenan
energía, lacual puede utilizarse para la conducción de otros
mecanismos de transporte activo. El gradienteelectroquímico
producido por estas bombas es tan importante que de hecho algunas
células (p. ej., lascélulas nerviosas) utilizan el 70% de su
energía en el funcionamiento de este sistema de transporte.
El uso de los potenciales electroquímicos con tales propósitos
no es exclusivo de la membranaplasmática de las células animales.
Las células de plantas y hongos también utilizan bombas
impulsadaspor ATP con las cuales "bombean" protones del citoplasma
hacia el exterior de la célula. La salida deprotones (con carga
positiva) del interior de la célula provoca una enorme diferencia
en la concentraciónde ellos, de manera que el exterior tiene carga
positiva y el interior negativo. Como se verá más adelante,estas
bombas de protones que utilizan ATP, usadas en sentido inverso para
producir ATP en bacterias,mitocondrias y cloroplastos, constituyen
el principal transductor de energía en todas las células, desde
lasbacterias hasta las células de plantas y animales complejos.
La hipótesis aceptada en la actualidad sostiene que la bomba de
sodio y potasio (y otras bombasque requieren ATP) está formada por
proteínas transmembranales que se extienden a través ella.
Laproteína tiene centros de unión para el Na+ y el ATP en su
superficie citoplasmática y para K + en susuperficie externa.
Después de una serie de cambios en su conformación son capaces de
intercambiarsodio por potasio a través de la membrana celular. A
diferencia de la difusión facilitada, parte de loscambios de
conformación de la proteína durante el ciclo de bombeo requieren de
la energía liberada por elATP. Parece que la energía se transfiere
del ATP a la bomba mediante la formación de un enlacecovalente
entre uno de los fosfatos del ATP y la proteína, seguido por la
retirada del mismo en etapas más
avanzadas del ciclo de bombeo. Debido al bombeo de 3 iones
Na+
positivamente cargados hacia elexterior de la célula cada dos
que bombea hacia el interior, se dice que la bomba es
electrogénica, esdecir dirige una corriente neta a través de la
membrana tendiendo a crear un potencial eléctrico, con el
-
8/17/2019 9 Membranas Biológicas y Transporte
15/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
15 15
interior negativo con relación al exterior. Por otra parte la
bomba tiene un papel directo regulando elvolumen celular a través
de sus efectos osmóticos, que pueden hacer que la célula se hinche
o seretraiga. Por último, también se utiliza para dirigir el
transporte de azúcares y aminoácidos hacia el interior
de la célula, como se verá a continuación.
Transporte activo secundario ocotransporte
Algunas proteínas de transporteactúan como transportadores
acoplados,en los que la transferencia de un solutodepende de la
transferencia simultánea osecuencial de un segundo soluto, ya seaen
la misma dirección (transporteunidireccional) o en dirección
opuesta
(bidireccional).En el sistema de transporte más simple, conocido
como uniporte, un soluto en particular se mueve
directamente a través de la membrana en una dirección. En el
tipo de cotransporte conocido comosimporte dos solutos diferentes
se mueven a través de la membrana, simultáneamente y en el
mismosentido. Así, las proteínas transportadoras actúan como
transportadores acoplados. Frecuentemente, ungradiente de
concentración, que involucra a uno de los solutos transportados,
impulsa el transporte delotro; por ejemplo, un gradiente de
concentración de iones Na+ frecuentemente impulsa el cotransporte
demoléculas de glucosa. En otro tipo de sistema de cotransporte,
conocido como antiporte, dos solutosdiferentes se mueven a través
de la membrana, simultánea o secuencialmente en sentidos opuestos.
Labomba o ATPasa Na+-K+ es un ejemplo de sistema de
cotransporte antiport o antiporte.
Sistemas de transporte múltiple integrado
En algunas células se observa el funcionamiento de más de un
sistema de transporte para unasustancia determinada. Por ejemplo,
el transporte de la glucosa del intestino hacia el torrente
circulatoriose lleva a cabo a través de una delgada capa de células
epiteliales que recubren la luz del intestino y queposeen regiones
especializadas, o dominios, en su membrana plasmática. La
superficie de estas células,expuesta en el intestino, posee una
gran cantidad de microvellosidades o protrusiones digitiformes
queincrementan con eficacia la superficie de membrana disponible
para absorción. El transporte de glucosa enesta zona de la
superficie celular es parte de un sistema de transporte activo que
se efectúa encotransporte con el sodio. La concentración
intracelular de sodio se mantiene baja por función de unabomba de
sodio y potasio en la superficie opuesta de la célula, que bombea
el sodio hacia el torrente
circulatorio. Gracias a su elevada concentración dentro de la
célula, la glucosa puede ser transportadahacia el torrente
circulatorio mediante difusión facilitada.
La localización de dos proteínas diferentes que participan en el
transporte de glucosa en dosregiones distintas de una misma
membrana plasmática se produce gracias a la presencia de
unionesespecializadas en la célula (“uniones estrechas”, se verá en
el módulo 6). Si una célula careciera de unmecanismo específico
para determinar este proceso, entonces las proteínas estarían
distribuidas al azaren ambas superficies de la célula; luego, no
habría transporte neto de glucosa.
-
8/17/2019 9 Membranas Biológicas y Transporte
16/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
16 16
Transporte de grandes moléculas a través de las membranas o
transporte en masa
En la difusión simple, en la difusión facilitada y en el
transporte activo las moléculas individuales ylos iones pasan a
través de la membrana celular. Sin embargo, en ocasiones también es
necesario eldesplazamiento de cantidades más grandes de material o
de partículas de alimento o incluso de célulascompletas, hacia
afuera o adentro de una célula. Esto implica un gasto de energía
por parte de la célula yen ocasiones conlleva también la fusión de
membranas. En la endocitosis, la célula incorpora materiales
hacia su interior. La endocitosis es el proceso celular, por el
que la célula mueve hacia su interiormoléculas grandes o
partículas, englobándolas en una invaginación de su membrana
citoplasmática,formando una vesícula que luego se desprende de la
membrana celular y se incorpora al citoplasma. Estavesícula,
llamada endosoma, luego se fusiona con un lisosoma que realizará la
digestión del contenidovesicular.
Existen tres procesos de endocitosis:1. Pinocitosis:
("bebido por células"), consiste en la ingestión de líquidos y
solutos mediante
pequeñas vesículas. En este tipo de endocitosis, la célula
incorpora materiales disueltos. Algunospliegues de la membrana
plasmática engloban gotas de líquido, las cuales emergen en
elcitoplasma celular en forma de pequeñas vesículas. El contenido
líquido de estas vesículas selibera lentamente en el citoplasma
celular y las vesículas van disminuyendo poco a poco de
tamaño, hasta el punto en que parecen desvanecerse.2.
Fagocitosis: (literalmente "ingesta de células") consiste en
la ingestión de grandes partículas
sólidas (como bacterias o nutrientes) que se engloban en grandes
vesículas (fagosomas) que se
La distribución asimétrica de las proteínas transportadoras en
la membrana plasmática de una célulaepitelial intestinal da lugar
al transporte transcelular de glucosa a través del epitelio
intestinal
-
8/17/2019 9 Membranas Biológicas y Transporte
17/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
17 17
desprenden de la membrana celular. La fagocitosis es el
mecanismo utilizado por protozoarios yleucocitos para ingerir
partículas, incluso algunas tan grandes como una bacteria
completa.Durante la ingestión, los pliegues de la membrana celular
engloban a la partícula, que se ha unido
a la superficie celular, y forman una vacuola o vesícula
alrededor de ella. Una vez que lamembrana ha encerrado a la
partícula en cuestión, se fusiona en el punto de contacto e
ingresaen el citoplasma. Posteriormente la membrana de la vesícula
se fusiona con la membrana dellisosoma (que contienen enzimas
hidrolíticas), donde el material es ingerido y degradado. A
vecestambién se degradan por este mecanismo organelos celulares
(una mitocondria, en el ejemplo),envolviendo al organelo en una
vesícula formada con membranas del retículo endoplásmico; aeste
proceso se lo denomina autofagocitosis.
3. Endocitosis mediada por receptor o ligando: es de
tipo específica, captura macromoléculas delambiente, fijándose a
través de proteínas receptoras ubicadas en la membrana plasmática.
Luego,las moléculas ligadas al receptor emigran hacia las
depresiones revestidas, que son regiones de
la superficie citoplásmica de la membrana recubiertas con
estructuras en forma de cepillo(proteínas denominadas clatrinas),
momentáneamente forman una estructura en forma de cestoalrededor de
ellas. Algunos segundos después, las vesículas son liberadas dentro
del citoplasma;sin embargo, el recubrimiento se separa de ellas,
dejando a las vesículas libres en él. En seguida,las vesículas se
fusionan con otras vesículas semejantes y forman endosomas,
vesículas másgrandes que transportan materiales libres, que no
están unidos a los receptores de membrana.Los endosomas forman dos
tipos de vesículas: unas contienen receptores que pueden regresar
ala membrana; otras, que contienen las partículas ingeridas, se
fusionan con los lisosomas ydespués son procesadas por la célula.
La endocitosis mediada por receptor resulta ser un procesorápido y
eficiente. La endocitosis mediada por receptores es un proceso
importante: por su medio,
Endocitosis mediada por receptores
-
8/17/2019 9 Membranas Biológicas y Transporte
18/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
18 18
las células animales incorporan hacia su interior el colesterol
del torrente circulatorio. Gran partedel mecanismo de esta variedad
de endocitosis fue detallada en estudios del receptor
delipoproteínas de baja densidad (LDL, “low density lipoproteins”).
El LDL está constituido por
proteína flanqueada por moléculas de fosfolípidos y colesterol;
en el centro hay un acumulo demoléculas de colesterol esterificado
con ácidos grasos de cadena larga. Las partículas de proteínade
baja densidad (LDL) se unen a proteínas receptoras específicas en
la membrana plasmática.Los complejos receptores de LDL se mueven a
lo largo de la superficie y se agrupan en la regiónde placas
recubiertas con clatrina de la membrana plasmática. La endocitosis
de tales placasrecubiertas da por resultado la formación de
vesículas recubiertas en el citoplasma celular.Algunos segundos
después el recubrimiento de clatrina se elimina y las vesículas se
fusionan conotras similares hasta formarse grandes vesículas
(endosomas jóvenes “early endosomes”). Enestas estructuras, los
receptores y las partículas de LDL se disocian, separan y dirigen
haciadiferentes regiones de la vesícula. A partir de los endosomas
se forman nuevas vesículas. Lasvesículas que contienen los
receptores se desplazan hacia la superficie y se fusionan con
la
membrana plasmática, en la cual los receptores son reciclados.
Las vesículas que contienenpartículas de LDL se fusionan con los
lisosomas. Finalmente el colesterol se libera por medio dediversas
enzimas hidrolíticas: sólo entonces puede utilizarlo la célula. Las
personas conincapacidad hereditaria para la producción de
receptores de membrana para el LDL (puede faltarel gen o fabricar
un receptor defectuoso, incapaz de unirse a la cubierta de
clatrinas) sonpropensas a arterioesclerosis prematura y a sufrir
ataques cardíacos.El reciclaje del receptor de LDL a la membrana
plasmática a través de la formación de vesículas
ilustra un problema común a todas las células que emplean los
mecanismos de endocitosis y exocitosis.En las células que secretan
sustancias en forma continua, unaporción equivalente de membrana
debe incorporarse al interiorde la célula por cada vesícula que se
fusiona con la membrana
plasmática; de no ocurrir así, la superficie de la célula
estaríaen expansión constante, aunque el crecimiento de la
célulaesté limitado. Existe una situación similar para las células
querealizan endocitosis. Los macrófagos, por ejemplo, incorporanen
forma de vesículas el equivalente del total de su volumen encerca
de 30 minutos, y requieren un reciclamiento similar paraque las
células mantengan su área de superficie y su volumen.
La exocitosis es el proceso celular por el cual
lasvesículas situadas en el citoplasma se fusionan con lamembrana
citoplasmática, liberando su contenido. En laexocitosis una
célula expulsa productos de desecho oproductos específicos de
secreción (como hormonas, p. ej.insulina), mediante la fusión de
una vesícula con la membranaplasmática de la célula. La exocitosis
consiste en la fusión de lamembrana de la vesícula secretora con la
membrana plasmática.Este es también un mecanismo primario de
crecimiento de lamembrana plasmática. La exocitosis se observa en
muy diversascélulas secretoras, tanto en la función de excreción
como en lafunción endocrina. También interviene la exocitosis en
lasecreción de un neurotransmisor a la brecha sináptica, para
posibilitar la propagación del impulso nervioso entre neuronas.
La secreción química desencadena unadespolarización del potencial
de membrana, desde el axón de
Neurona A (transmisora) a neurona B (receptora)
1. Mitocondria2. Vesícula sináptica
con neurotransmisores
3. Autoreceptor
4. Sinapsis con neurotransmisores liberados
(Serotonina)
5. Receptores Post-sinápticos activados por
neurotransmisores (inducción de
un Potencial postsináptico)
6. Canal de calcio
7. Exocitosis de una vesícula
8. neurotransmisor recapturado.
http://es.wikipedia.org/wiki/Mitocondriahttp://es.wikipedia.org/wiki/Ves%C3%ADcula_sin%C3%A1pticahttp://es.wikipedia.org/wiki/Neurotransmisorhttp://es.wikipedia.org/wiki/Sinapsishttp://es.wikipedia.org/wiki/Serotoninahttp://es.wikipedia.org/wiki/Serotoninahttp://es.wikipedia.org/w/index.php?title=Potencial_postsin%C3%A1ptico&action=edit&redlink=1http://es.wikipedia.org/w/index.php?title=Potencial_postsin%C3%A1ptico&action=edit&redlink=1http://es.wikipedia.org/wiki/Canal_de_calciohttp://es.wikipedia.org/wiki/Canal_de_calciohttp://es.wikipedia.org/w/index.php?title=Potencial_postsin%C3%A1ptico&action=edit&redlink=1http://es.wikipedia.org/w/index.php?title=Potencial_postsin%C3%A1ptico&action=edit&redlink=1http://es.wikipedia.org/wiki/Serotoninahttp://es.wikipedia.org/wiki/Sinapsishttp://es.wikipedia.org/wiki/Neurotransmisorhttp://es.wikipedia.org/wiki/Ves%C3%ADcula_sin%C3%A1pticahttp://es.wikipedia.org/wiki/Mitocondria
-
8/17/2019 9 Membranas Biológicas y Transporte
19/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
19 19
la célula emisora hacia la dendrita (u otra parte) de la célula
receptora. Este neurotransmisor será luegorecuperado por
endocitosis para ser reutilizado. Sin este proceso, se produciríaun
fracaso en latransmisión del impulso nervioso entre neuronas.
La secreción puede ser constitutiva, como cierto tipo de
proteínas que son regularmentesecretadas por la célula, o ser una
secreción regulada, cuando es necesario que una sustancia
externa(una señal, como una hormona o un neurotransmisor)
desencadene un proceso (transducción de señal)que finaliza con la
secreción de una sustancia producida por la célula que es requerida
en el medioextracelular.
-
8/17/2019 9 Membranas Biológicas y Transporte
20/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
20 20
BIBLIOGRAFIA
Alberts B., D. Bray, J. Lewis., M. Raff, K. Roberts & J.D.
Watson. Biología Molecular de la Célula. 3ra Ed.
1996.Cooper, G.M. (2002). La Célula. 2ª edición. Marbán Libros,
S.L., España. (Traducido de la 2ª edicióninglesa, 2000).
Curtis, H. y N.S. Barnes (2000) “Biología”. 6ª edición española.
Editorial Médica Panamericana. Lehninger, A.L., D.L. Nelson y
M.M. Cox (1993) “Principios de Bioquímica”, Ediciones Omega,
Barcelona, 2a
edición (traducido de la segunda edición inglesa. 1993).Lodish,
H., A. Berk, S.L. Zipursky, P. Matsudaira, D. Baltimore & J.
Darnell (2002)”Biología Celular y
Molecular”, Editorial Médica Panamericana (traducido de la 4ª.
Ed. inglesa, 2000). Purves, K.W., D. Sadava, G.H. Orians &
H.C. Heller (2003) “Vida. La Ciencia de la Biología”, 6ª.
Edición.
Editorial Médica Panamericana (traducido de la 6ª edición
inglesa, 2001).Solomon, E.P., L. R. Berg, D.W. Martin & C.A.
Villee (1996) “La Biología de Villee”, Interamericana
McGraw-Hill.Stryer L.(1992). Bioquímica. Ed. Reverté. 3ra
Edic.1992.
CUESTIONARIO TEÓRICO1. ¿Por qué se dice que las membranas
celulares no son paredes rígidas sino estructuras complejas y
dinámicas?2. ¿Cómo podría explicar el “modelo del mosaico
fluido” y cuál es su relación con el comportamiento de las
membranas biológicas?3. ¿Qué características tiene una
sustancia anfipática? ¿Todas las moléculas que integran la
membrana
plasmática son anfipáticas?
4. ¿La membrana plasmática es simétrica o asimétrica?
¿Cuál es el efecto funcional de este hecho?5. ¿Cuál es la
función de las proteínas presentes en la membrana plasmática? ¿Qué
tipos de proteínas de
membrana conoce?6. ¿Cómo explica que si en la membrana
plasmática hay un 50% de lípidos y otro tanto de proteínas, la
relación numérica sea de alrededor de 50 moléculas de
fosfolípidos por cada molécula de proteína?7. Explique por
qué es falsa esta expresión: “la diferencia entre proteínas
solubles y proteínas ligadas a
membranas consiste en que una no contiene aminoácidos
hidrofóbicos y la otra sí” 8. ¿Qué diferencias existen
entre los fenómenos de difusión, diálisis y ósmosis?9. ¿De
qué depende que una solución sea isotónica, hipotónica o
hipertónica?10. Los iones, azúcares, aminoácidos, nucleótidos
y muchos metabolitos ingresan a la célula por el
mecanismo de difusión facilitada. ¿Es necesario algún componente
especial en la membranaplasmática para que esto suceda?
11. ¿A qué se denomina “transporte activo” de moléculas a
través de la membrana plasmática? Mencioneejemplos.
12. ¿Qué diferencia hay entre la endocitosis, la
fagocitosis y la autofagia? ¿Qué es lo que tienen en
común?13. Describa la endocitosis mediada por
receptores.14. ¿En qué consiste un sistema de transporte
múltiple integrado? Proporcione un ejemplo.15. ¿Qué entiende
por exocitosis? ¿Qué diferencias existen entre una secreción
constitutiva y una secreción
regulada?
-
8/17/2019 9 Membranas Biológicas y Transporte
21/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
21 21
El transportador defectuoso de la fibrosis quística
UN ACERCAMIENTO A LAS MEMBRANAS
CELULARESSin importar qué tan pequeña sea una célula, esun ser
vivo que lleva a cabo operacionespeligrosas. Piensa en las células
de todo tu cuerpoy en cómo deben responder a algo tan comúncomo el
agua. En todo momento deben desplazarel agua y los solutos en uno y
otro sentido de sumembrana plasmática. Las sustancias que pasana
través de la membrana deben sercuidadosamente seleccionadas, ya que
delmantenimiento de las condiciones favorablesdepende la
supervivencia de cada célula, en último
término, de todo el cuerpo. Cuando todo va bien,las células
toman y expulsan las cantidadesprecisas, ni muy bajas, ni
excesivas. Pero nosiempre salen bien las cosas. Existe un
canalproteico, llamado CFTR, que como todos loscanales atraviesa la
membrana plasmática de lascélulas epiteliales. Este tipo de células
formado untejido formado por una o varias capas de célulasunidas
entre si que recubren todas las superficieslibres del organismo, y
constituyen el recubrimientointerno de las cavidades, órganos
huecos,conductos del cuerpo y la piel y que también
forman las mucosas y las glándulas.
Representación esquemática del canal CFTR. Este secompone de 5
dominios: dos dominios membrana-llave,que forman el canal del ion
cloruro; dos dominiosnucleótidos-vinculantes que unen e hidrolizan
ATP, y undominio regulador.
En condiciones de salud, los iones cloruroatraviesan dichas
células junto con el agua para
formar una delgada capa sobre la superficie deestos
recubrimientos. La mucosidad, que lubrica
los tejidos y ayuda a evitar infecciones, se deslizalibremente y
sin dificultad sobre la película acuosa.
En ocasiones, la codificación genética para elregulador de la
conductancia transmembrana de lafibrosis quística, en inglés cystic
fibrosistransmembrane conductance regulator o para losamigos CFTR,
muta lo que provoca que el cloruroy el agua no atraviesen de manera
adecuada losepitelios, de modo que no se forma el
resbaladizorecubrimiento. En su ausencia, la mucosidad sehace
espesa, se seca y su flujo se dificulta. Estoprovoca que, por
ejemplo, se obstruyan losconductos del páncreas, que las
enzimas
digestivas no puedan llegar al intestino delgado, enconsecuencia
los alimentos no se digieren y noson absorbidos lo que genera a su
vez que lapersona baje de peso. Las glándulas sudoríparasexcretan
el exceso de sal alterando el equilibrio deagua y sodio del cuerpo,
lo cual afecta al corazón ya todos los órganos con tejido
epitelial. En losvarones ocasiona esterilidad.
¿Pero qué ocurre con las vías respiratorias? Por lo regular,las
células ciliadas, con finas estructuras semejantes apelos, barren
los gérmenes y partículas de polvo que
inhalamos y que son atrapadas en la capa de moco querecubre las
vías aéreas. Pero en este caso no sucede así,se forman biocapas,
estructuras espesas y pegajosasconstituidas de poblaciones de
bacterias ancladas sobre elrecubrimiento mucoso. Las biocapas
resisten fuertementelas defensas del cuerpo y a los antibióticos.
LaPseudomona aeruginosa es la más eficiente de
lascolonizadoras, y es la causante de infecciones crónicas.
Lamayoría de los pacientes con fibrosis quística (FQ) muerenpor
insuficiencia pulmonar y, en general no tienen unaexpectativa de
vida mayor de 30 años.
La CFTR mutante es la proteína que inicia todos estoseventos de
la FQ, que en la actualidad es uno de lostrastornos genéticos
mortales más común en niños. De
-
8/17/2019 9 Membranas Biológicas y Transporte
22/22
CÉLULA I/Membranas Biológicas y TransporteGonzalo Vázquez
Palacios
22 22
acuerdo con la AsociaciónMexicana de FQ, alrededor de400 niños
nacen cada año en
México con esta enfermedad, sin embargo, con losrecursos e
infraestructura actuales, solamente el 15% deellos son
diagnosticados con vida, el resto, 85%, falleceantes de cumplir los
4 años de edad por complicacionesrespiratorias y desnutrición. Se
cree que existenactualmente en nuestro país alrededor de 6,000
niños conFQ los cuales no tienen acceso a un
tratamientoespecializado.
La CFTR pertenece a la familia de transportadores ABC,que son
proteínas membranales presentes en todas lasespecies procariontes y
eucariontes. El gen para CFTR,cuya alteración es responsable de
esta enfermedad, tiene27 exones y se extiende sobre 250 kb del
cromosoma 7(7q31) dando lugar a un RNAm de 6,5 kb. La mutación
máscomún es una deleción de los tres nucleótidos que
componen al codón de la fenilalanina (F) en la posición 508del
gen. Una persona con la mutación CFTRΔF508producirá la proteína
CFTR que carece de este residuo defenilalanina, por lo que no
funciona correctamente. En el70% de los pacientes con FQ, la
pérdida de un soloaminoácido durante la síntesis de la proteína
provoca estetrastorno mortal. Antes de que la CFTR sea llevada a
lamembrana celular, se cree que es modificada por elsistema
endomembranal. Copias de la proteína mutanteentran al retículo
endoplásmico, pero las enzimas no lareconocen y destruyen el 99% de
las mismas antes de quelleguen al aparato de Golgi. De este modo
muy pocos
canales de CFTR llegan a su destino. Este canal
tambiéncontribuye a problemas de sinusitis, esto es unainflamación
crónica del recubrimiento de las cavidades delcráneo cercanas a la
nariz.Con este ejemplo se puede ver cómo surgen problemas desalud
cuando las células no tienen copias correctas de tansólo una de las
proteínas de la membrana plasmática.Piénsalo un poco y podrás
apreciar la precisión con la quela célula utiliza sus proteínas.
Esta precisión también seextiende a las membranas de los
compartimientoscelulares internos llamados organelos. Al comprender
laestructura de las membranas celulares podrás conocer
también su funcionamiento y cómo se efectúa lasupervivencia al
nivel fundamental de la vida.Para saber más:
1) Fibrosis quística: la frontera del conocimiento
molecular y susaplicaciones clínicas. Lorena Orozco, et al. (2006).
Rev InvClin 58(2):
139-152.http://www.scielo.org.mx/pdf/ric/v58n2/v58n2a7.pdf
2) Fibrosis quística: Guías clínicas para el diagnostico
ytratamiento, Dr. José Luis Lezama Fernández (Editor).
Editorial Intersistemas. México D.F.,
2008.http://www.fibrosisquistica.org.mx/home/data/docs/Fibrosis_Quistica.pdf
3) La fibrosis quística y el gen
CFTR.http://atlasgeneticsoncology.org/Educ/CistFibID30032SS.html
4) CFTR: Más que un canal de cloro. Dr. Luis E.
Vega-Briceño.Rev Chil Pediatr 2005;
76:464-70.http://www.neumologia-pediatrica.cl/pdf/201051/CFTR.pdf
5) CFTR: 15 años después del descubrimiento de un
gen.VEGA-BRICEÑO Luis Enrique. Rev Med Hered 15 (3), 2004159.
http://www.scielo.org.pe/pdf/rmh/v15n3/v15n3tr1.pdfAsociación
Mexicana de Fibrosis Quística A.C.
(AMFQ)http://www.fibrosisquistica.org.mx/home/index.php?id=1
http://www.scielo.org.mx/pdf/ric/v58n2/v58n2a7.pdfhttp://www.scielo.org.mx/pdf/ric/v58n2/v58n2a7.pdfhttp://www.fibrosisquistica.org.mx/home/data/docs/Fibrosis_Quistica.pdfhttp://www.fibrosisquistica.org.mx/home/data/docs/Fibrosis_Quistica.pdfhttp://www.fibrosisquistica.org.mx/home/data/docs/Fibrosis_Quistica.pdfhttp://atlasgeneticsoncology.org/Educ/CistFibID30032SS.htmlhttp://atlasgeneticsoncology.org/Educ/CistFibID30032SS.htmlhttp://www.neumologia-pediatrica.cl/pdf/201051/CFTR.pdfhttp://www.neumologia-pediatrica.cl/pdf/201051/CFTR.pdfhttp://www.fibrosisquistica.org.mx/home/index.php?id=1http://www.fibrosisquistica.org.mx/home/index.php?id=1http://www.fibrosisquistica.org.mx/home/index.php?id=1http://www.neumologia-pediatrica.cl/pdf/201051/CFTR.pdfhttp://atlasgeneticsoncology.org/Educ/CistFibID30032SS.htmlhttp://www.fibrosisquistica.org.mx/home/data/docs/Fibrosis_Quistica.pdfhttp://www.fibrosisquistica.org.mx/home/data/docs/Fibrosis_Quistica.pdfhttp://www.scielo.org.mx/pdf/ric/v58n2/v58n2a7.pdf