Embed Size (px)
Citation preview

QUATERNARY RESEARCH 21, 225-243 (1984)
13C Record of Benthic Foraminifera in the Last Interglacial Ocean: Implications for the Carbon Cycle and the Global Deep
Water Circulation
JEAN-CLAUDE DUPLESSY,* NICHOLAS J. SHACKLETON.? ROBLEY K. MATTHEWSA WARREN PRELL,$. WILLIAM F. RUDDIMAN,§ MICHBLECARALP,~~ AND
CHRIS H. HENDY*,~
* CLIMAP, Centre des Faibles RadioactivitPs, Laboratoire mixte CNRS-CEA, 91190 Gif sur Yvette, France; tCLIMAP, The Godwin Laboratory, Free School Lane, Cambridge CB2 3RS, Great Britain; SCLIMAP, Department of Geological Sciences, Brown University, Providence, Rhode Island 02912:
KLIMAP, Lamont Doherty Geological Observatory, Palisades, New York 10964; and IlLaboratoire de Ge’ologie et Oc&anographie, Universite de Bordeaux I, Avenue des Facult&, 33405 Talence, France
Received February 14, 1983
The t3C/t2C ratios of Upper Holocene benthic foraminiferal tests (genera Cibicides and Uvi- gerina) of deep sea cores from the various world ocean basins have been compared with those of the modern total carbon dioxide (TCO?) measured during the GEOSECS program. The 6r3C dif- ference between benthic foraminifera and TCO, is 0.07 ? 0.04%0 for Cibicides and -0.83 -+ 0.07%c for Uvigerina at the 95% confidence level. 6t3C analyses of the benthic foraminifera that lived during the last interglaciation (isotopic substage Se, about 120,000 yr ago) show that the bulk of the TCOz in the world ocean had a 613C value 0.15 ? 0.12%~ lower than the modem one at the 95% confidence level, reflecting a depletion. compared to the present value, of the global organic carbon reservoir. Regional differences in 6r3C between the various oceanic basins are explained by a pattern of deep water circulation different from the modem one: the Antarctic Bottom Water production was higher than today during the last interglaciation. but the eastward transport in the Circumpolar Deep Water was lower.
INTRODUCTION
Deep water is formed at high latitudes, where the atmosphere extracts heat from the sea surface and makes the surface water sufficiently dense to sink to great depth and flow toward the equator. This formation process is dependent on the climate, so that the formation and circulation of oceanic deep water are affected by the climatic changes that the earth underwent during the Quaternary. Streeter (1973), Schnitker (1974), Lohmann (1978), and Streeter and Shackleton (1979) have shown that the rel- ative abundance of benthic foraminiferal taxa varies within the Pleistocene with striking changes occurring within a few thousand years. These variations have been
r Present address: University of Waikato, Chem- istry Department, Hamilton, New Zealand.
explained in terms of changes either in the origin or in the oxygen content of the deep water masses.
Oxygen isotope variations in benthic for- aminifera from deep-sea cores have also been used to reconstruct some trends of deep water paleocirculation. Duplessy er al. (1975) demonstrated that during the last glacial age, Norwegian Sea Deep Water was warmer than at present. This result indi- cates that the Norwegian Sea was not a sink for surface water, as a result of the disap- pearance of the Norwegian current (Kel- logg, 1976) and stratification due to a per- manent ice cover (Kellogg et al., 1978). This implies a major change in the origin and circulation of the deep water of the North Atlantic Ocean since, under modern conditions, the Norwegian Sea is the major source of North Atlantic Deep Water
225 0033-5894184 $3.00 Copyright 8 1984 by the Univemry of Washington.
*..s.t. -c rr*r”Al.ni.... i.. -.../ carm r~c~r”r,i

(NADW; Worthington, 1976). Subse- the total CO2 dissolved in the deep ocean quently, Duplessy et al. (1980) showed that becomes progressively lighter during the the oxygen isotopic composition of benthic transit of the deep water mass from its foraminifera from the northeastern Atlantic source in the North Atlantic Ocean to the Ocean was richer in 180 than that of benthic other deep oceanic basins. As a conse- foraminifera from the southern Indian quence, the highest 13C content for TCO, is Ocean during the last glaciation. This dif- at present found in the Norwegian Sea and ference was interpreted as an indication in the North Atlantic Ocean (Kroopnick, that glacial NADW was cooler than it is 1980) whereas the lowest values come from today by at least 1.3”C and that this water the Pacific (Kroopnick, 1974). This is a re- did not originate from the Norwegian Sea sult of the larger oxygen utilization and of overflow, but from the sinking of surface the longer residence time (a factor of three) water in the North Atlantic Ocean. of the deep water in the Pacific than in the
Neither benthic fauna analysis nor the Atlantic. comparison of 180 content of benthic fora- We first show that, as a first approxi- minifera from various oceanic basins has mation, the carbon isotope ratio of the fo- proved useful in reconstructing the major raminiferal genus Cibicides directly reflects trends of the deep water circulation in the that of TCO, whereas the genus Uvigerina world ocean. This is mainly due to the high exhibits a departure from this value which degree of intercorrelation that exists in the can be assumed constant. We then use the deep sea between the various environ- t3C analyses of the benthic foraminifera mental properties in any one area, which that lived during the last interglaciation to makes it difficult to recognize the factors reconstruct the 13C gradient between the determining the abundance of a foraminif- various oceanic basins and to deduce the era1 species (Schnitker, 1980). Concerning major trends of the deep water circulation the oxygen isotope method, the small range at that time. of temperature variations from one basin to The four isotope laboratories involved in another produces only minor F180 varia- isotope studies for the CLIMAP Project tions, which are difficult to extract from the (Brown University, Cambridge, Gif sur experimental noise. Yvette, and Lamont Doherty Geological
In this paper, we attempt to use the Observatory) use an experimental proce- t3C/12C ratio of benthic foraminifera to com- dure derived from one set up by Shackleton pare the deep water circulation in the last and described by Shackleton and Opdyke interglacial ocean with that of the modern (1973, 1976) and Duplessy (1978). These ocean. The rationale behind this approach laboratories were intercalibrated with the is the recent conclusion that the dominant same working standards, allowing compar- factor affecting the distribution of 6t3C of ison of their results with minimal error. the total dissolved CO, (TCO,) in the world ocean is the deep water circulation (Craig, RELIABILITY OF THE BENTHIC
1970; Kroopnick et al., 1970; Duplessy, CARBON ISOTOPE RECORD
1972; Kroopnick, 1974, 1980). In the deep In order to infer past oceanographic con- ocean, oxygen is consumed by the respi- ditions from the fossil isotope record, we ration of living organisms and by the de- must determine how accurately the upper composition of organic material by bacteria Holocene benthic foraminifera reflect the (Richards, 1965) and it is this oxygen con- conditions prevailing today in the deep sumption that releases CO, in the deep ocean. The oxygen isotope composition of ocean. Because the organic matter is de- benthic foraminifera has been shown to re- pleted in t3C by some 20 to 25%0 (Sackett et flect the deep water temperature and iso- al., 1965; Fontugne and Duplessy, 1981), tope ratio, after correction for a constant,
226 DUPLESSY ET AL.

INTERGLACIAL OCEANIC 13C RECORD 227
species-dependent, departure from isotopic equilibrium (Duplessy et al., 1970; Shack- leton and Opdyke, 1973; Woodruff et al., 1980; Belanger et al., 1981; Vincent et al., 1981a). The reliability of the carbon isotope record of benthic foraminifera is much less well established.
Previous Work
Shackleton (1977) was the first to explore systematically the 13C record in benthic fora- minifera. He found that their 6r3C values were always lower during the last ice age than during the Holocene and he inter- preted this change as the result of de- creased forest cover during glacial time. Subsequently, the CENOP group (see a synthesis by Savin et al. (1981)) recognized a late Miocene carbon shift reflecting an ap- parently synchronous world-wide decrease in foraminiferal 6r3C, which must have re- sulted from a simultaneous decrease in 613C of the total CO, dissolved in the whole ocean. Consequently, the occurrence of well-defined signals in carbon isotope rec- ord of fossil benthic foraminifera seems well established.
Boersma and Shackleton (1977) used 13C differences between benthic foraminifera in two DSDP sites (366 and 357) to show that during Oligocene time, deep water in the equatorial Atlantic was depleted in dis- solved oxygen by about 2 ml/liter compared with the South Atlantic. More recently, Shackleton et al. (1983) have made a detailed comparison between an eastern Atlantic and a Pacific late Pleistocene 13C record to show that deep water in both oceanic basins has similar age during glacial conditions.
Recently Belanger et al. (1981) and Graham er al. (1981) have shown that de- spite species-dependent degrees of 13C dis- equilibrium, the benthic foraminifera from core-top sediment correctly reflect the ob- served carbon isotope gradient between the Atlantic and Pacific oceans. Curry and Lohmann (1982) used 613C values of Cibi-
tides wuellestorfi to reconstruct the av- erage bathymetric profile of 613C during glacial stages in the Vema Channel and to deduce the stratification of deep water masses under glacial conditions. Finally, Blanc (1981) and Blanc and Duplessy (1983) compared benthic records from Atlantic and Pacific DSDP sites to reconstruct deep water circulation from middle Miocene times to the Pliocene and to show the im- pact of the Messinian salinity crisis on the formation of the North Atlantic Deep Water.
Calibration of Cibicides and Uvigerinu against Total Dissolved Carbon Dioxide 613C
Generally any single benthic foraminif- era1 species is not abundant enough for analysis throughout the length of a deep-sea core. In order to get a complete isotopic record, one reference species is chosen and the other species are compared to that ref- erence species. The final isotope record is pieced together from the analysis of several taxa after adjustments have been made to compensate for isotopic departure of the various taxa from the reference species. The isotope departure between different species is estimated by comparing the iso- topic composition of the various taxa when they are sufficiently abundant at the same core level.
Because the cores used for the recon- struction of the last interglacial ocean have been sampled in far greater detail than is customary in deep-sea core studies, the preceding procedure for deriving adjust- ments between different species appeared to be possibly misleading. As an example, in the northeastern Atlantic Ocean, Uvi- gerinu peregrinu is the dominant benthic species during isotope stage 6 and isotope substage 5d, whereas Plunulinu wuelle- storJi is absent. By contrast this last species becomes dominant during the last intergla- cial stage 5e when U. peregrinu is absent (Streeter and Shackleton, 1979). Hence, U. peregrinu cannot be calibrated against P. wuellestorfi because these two species may

228 DUPLESSY ET AL.
have never cohabited. In some sediment levels, generally corresponding to climatic transitions, both species can be found to- gether. In fact, this co-occurrence primarily corresponds to individuals shifted by bio- turbation upward or downward from their original level in the core (Peng et al., 1977; Duplessy, 1978; Hutson, 1980). If these samples were used for calibration, they would lead to wrong adjustments since a glacial sample of a given species would be compared to an interglacial sample of an- other species.
In order to avoid this difficulty, we only analyzed benthic foraminifera from upper Holocene sediment. We used cores for which an oxygen isotope stratigraphy was previously established and a few additional samples for which the 6180 values were used to check that the age is recent. Core locations are shown in Figure 1 and re- ported in Table 1. We calibrated Uvigerina and Cibicides by comparing their measured 613C values with that of the total CO, dis-
solved in the sea water at the location and depth of the core. This was a feasible pro- cedure because the 13C distribution in the Atlantic, Indian, and Southern oceans is well established (Kroopnick, 198 1; unpub- lished GEOSECS data). The coverage is far more scarce in the Pacific Ocean because GEOSECS data are not yet available. Nev- ertheless the data of Craig (1970), Kroop- nick et al. (1972), and Kroopnick (1974) have been used to estimate the 613C of TCO, with an accuracy which is >0.1%0 when the cores are far from a hydrologic station. Fortunately many cores were col- lected in the equatorial Pacific and in the Panama Basin, where reliable 13C data have been published by Kroopnick (1974), pro- viding more accurate S13C estimates.
Analytical values for 613C of TCO, under modern conditions and for 613C of C. wuel- lestor- and Uvigerina sp. from upper Ho- locene sediment are reported in Table 2. We have also included a few analyses of Cibi- tides kullenbergi and Cibicides pseudoun-
70. rn’ 30’ Ed so’ ml’ Ed 190’ Isd 120’ 90’ 60’ 30. 0' 30'
FIG. 1. Location of cores used either for the calibration of the benthic foraminifera Cibicides and Uvigerina or for the reconstruction of S’3CTco, in the last interglacial ocean.
INTERGLACIAL OCEANIC 13C RECORD 229
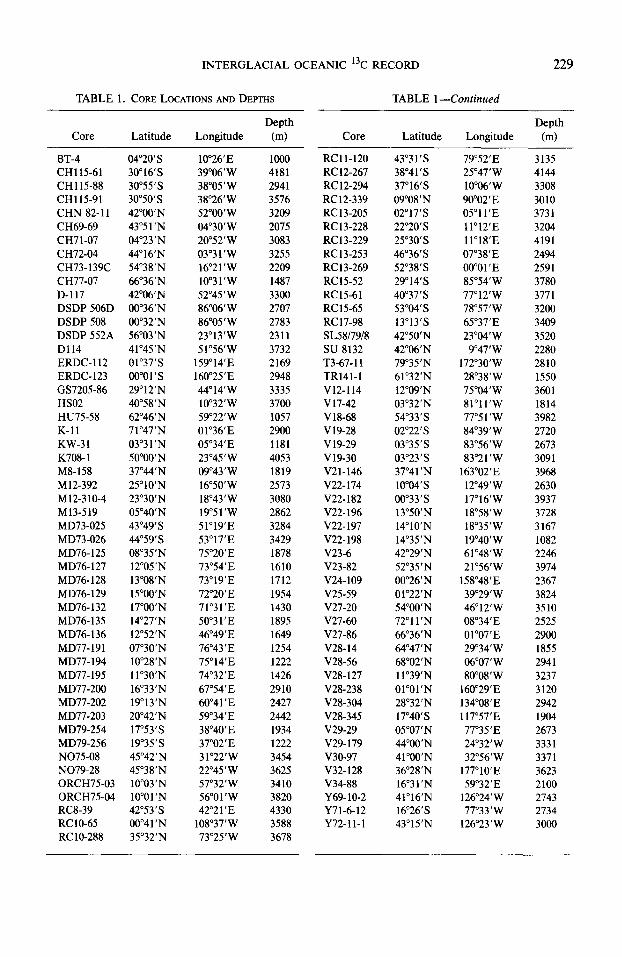
TABLE 1. CORE LOCATIONS AND DEWS TABLE 1 -Continued
Core Latitude Depth
Longitude (m) Core Latitude Longitude
BT-4 04”2O’S lO”26’E 1000 CHllS-61 30”16’S 39”06’W 4181 CH115-88 30”55’S 38”05’W 2941 CHl15-91 30”5O’S 38”26’W 3576 CHN 82-11 42”OO’N 52”OO’W 3209 CH69-69 43’51’N 04”3O’W 2075 CH71-07 04”23’N 20’52’W 3083 CH72-04 44”16’N 03”3 1 ‘W 3255 CH73-139C 54”38’N 16”21’W 2209 CH77-07 66’36’N lO”31’W 1487 D-117 42’06’N 52”45’W 3300 DSDP 506D OO”36’N 86”06’W 2707 DSDP 508 00”32’N 86’05 ’ W 2783 DSDP 552A 56’03’N 23’13’W 2311 D114 41’45’N 51”56’W 3732 ERDC-112 Ol”37’S 159”14’E 2169 ERDC- 123 oo”ol’s 160’25’E 2948 GS7205-86 29’12’N 44”14’W 3335 HS02 40’58’N lO”32’W 3700 HU75-58 62”46’N 59”22’W 1057 K-11 71’47’N Ol”36’E 2900 KW-31 03’3 1 ‘N 05’34’E 1181 K708-1 50”OO’N 23’45’W 4053 MS-158 37”44’N 09”43’W 1819 M12-392 25”lO’N 16’5O’W 2573 M12-310-4 23’30’N 18’43’W 3080 M13-519 OS”40’N 19”51’W 2862 MD73-025 43”49’S 51’19’E 3284 MD73-026 44”59’S 53’17’E 3429 MD76-125 OSgS’N 75”20’E 1878 MD76-127 12”OS’N 73’54’E 1610 MD76-128 13”08’N 73’19’E 1712 MD76-129 15”OO’N 72”20’E 1954 MD76-132 17”OO’N 71’31’E 1430 MD76-135 14”27’N 50”31’E 1895 MD76- 136 12”52’N 46”49’E 1649 MD77-191 07”30’N 76”43’E 1254 MD77-194 lO”28’N 75”14’E 1222 MD77- 195 1 l”30’N 74”32’E 1426 MD77-200 16’33’N 67’54’E 2910 MD77-202 19”13’N 60”41’E 2427 MD77-203 20”42’N 59”34’E 2442 MD79-254 17”53’S 38’40’E 1934 MD79-256 19”35’S 37”02’E 1222 N075-08 45”42’N 31’22’W 3454 N079-28 45’38’N 22’45’W 3625 ORCH75-03 lO”03’N 57’32’W 3410 ORCH75-04 10”Ol’N 56”Ol’W 3820 RCS-39 42’53’s 42’21 ‘E 4330 RClO-65 OO”41’N lOs”37’W 3588 RC lo-288 35”32’N 73’25’W 3678
RCll-120 RC 12-267 RC 12-294 RC12-339 RC13-205 RC13-228 RC13-229 RC13-253 RC13-269 RC15-52 RC15-61 RC15-65 RC17-98 SL58/79/8 SU 8132 T3-67- 11 TR141-1 V12-114 V17-42 V18-68 V 19-28 Vl9-29 v19-30 V21-146 V22-174 V22-182 V22-196 V22-197 V22-198 V23-6 V23-82 V24-109 V25-59 V27-20 V27-60 V27-86 V28-14 V28-56 V28-127 V28-238 V28-304 V28-345 V29-29 V29-179 v30-97 V32-128 V34-88 Y69-10-2 Y71-6-12 Y72-1 l-l
43”31’S 79”52’E 38”41’S 25”47’W 37=16’S lO”O6’W 09’08’N 90”02’E 02”17’S 05”ll’E 22”2O’S 1 l”12’E 25’30’s 1 l”l8’E 46”36’S 07”38’E 52’38’s 00”Ol’E 29”14’S 85”54’W 40”37’S 77”12’W 53”04’S 78”57’W 13”13’S 65”37’E 42”50’N 23’04’W 42”06’N 9”47’W 79”35’N 172”3O’W 61’32’N 28”38’W 12”W’N 75”04’W 03”32’N 81’1 I’W 5433’S 77”5l’W 02”22’S 84’39’W 03”35’S 83’56’W 03’23’s 83’21’W 37”41 ‘N 163”02’E lO”o4’S 12’49’W OO”33’S 17’16’W 13’50’N 18”58’W 14”lO’N 18”35’W 14”35’N 19”4O’W 42”29’N 61’48’W 52”35’N 21’56’W OO”26’N 158”48’E Ol”22’N 39’29’W 54”OO’N 46”12’W 72”ll’N 08”34’E 66”36’N Ol”07’E 64’47’N 29”34’W 68”02’N 06’07’W 1 l”39’N 8o”OS’W 0l”Ol’N 160”29’E 28”32’N 134’08’E 17”4O’S 117’57’E 05”07’N 77”35’E 44”OO’N 24’32’W 41”OO’N 32”56’W 36”28’N 177”lO’E 16’31’N 59’32’E 41”16’N 126”24’W 16’26’s 77”33’W 43’15’N 126’23’W
-
Depth
6-4
3135 4144 3308 3010 3731 3204 4191 2494 2591 3780 3771 3200 3409 3520 2280 2810 1550 3601 1814 3982 2720 2673 3091 3968 2630 3937 3728 3167 1082 2246 3974 2367 3824 3510 2525 2900 1855 2941 3237 3120 2942 1904 2673 3331 3371 3623 2100 2743 2734 3000
230 DUPLESSY ET AL.
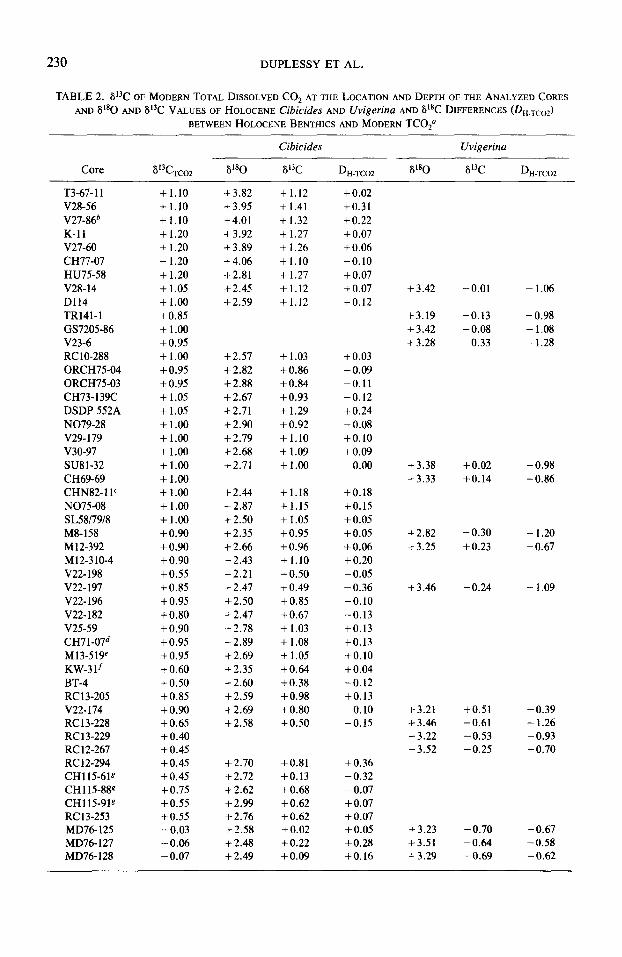
TABLE2. 613C OFMODERNTOTALDISSOLVEDCO~ AT THE LOCATION AND DEPTHOFTHEANALYZEDCORES AND 8'*0 AND 813C VALUES OF HOLOCENE Cibicides AND Uvigerina AND 6’8C DIFFERENCES (DH.TCOZ)
BETWEEN HOLOCENE BENTHICS AND MODERN TCOza
Core
T3-67-11 V28-56 V27-86b K-11 V27-60 CH77-07 HU75-58 V28-14 D114 TR141-1 GS7205-86 V23-6 RC lo-288 ORCH75-04 ORCH75-03 CH73-139C DSDP 552A N079-28 V29-179 v30-97 SU81-32 CH69-69 CHN82- 1 lc N075-08 SL5817918 M8-158 M12-392 M12-310-4 V22-198 V22-197 v22-1% V22- 182 V25-59 CH71-07d M13-519e KW-3 lf BT-4 RC13-205 V22-174 RC13-228 RC13-229 RC 12-267 RC 12-294 CH115-61g CH115-889 CH115-919 RC13-253 MD76- 125 MD76-127 MD76-128
-
Cibicides Uvigerina
6’3C TCO2 6’8O 8i3C D H-TCO2 8’8O iY3C D~-~co2
+1.10 + 1.10 +1.10 +1.20 +1.20 + 1.20 + 1.20 + 1.05 +1.00 +0.85 + 1.00 +0.95 +1.00 +0.95 +0.95 + 1.05 + 1.05 + 1.00 + 1.00 + 1.00 + 1.00 + 1.00 + 1.00 +1.00 +1.00 +0.90 +0.90 +0.90 +0.55 +0.85 + 0.95 +0.80 +0.90 +0.95 +0.95 + 0.60 + 0.50 + 0.85 +0.90 +0.65 +0.40 +0.45 +0.45 + 0.45 +0.75 +0.55 +0.55 -0.03 -0.06 -0.07
+3.82 +1.12 +3.95 + 1.41 +4.01 + 1.32 +3.92 + 1.27 + 3.89 +1.26 + 4.06 +1.10 +2.81 +1.27 +2.45 +1.12 +2.59 +1.12
d-2.57 +2.82 +2.88 + 2.67 +2.71 +2.90 + 2.79 + 2.68 +2.71
+1.03 + 0.86 + 0.84 +0.93 + 1.29 + 0.92 +1.10 +1.09 +1.00
+2.44 + 2.87 +2.50 + 2.35 f2.66 +2.43 +2.21 f2.47 +2.50 + 2.47 i-2.78 +2.89 f2.69 + 2.35 + 2.60 + 2.59 + 2.69 +2.58
+1.18 +1.15 +1.05 +0.95 +0.96 + 1.10 + 0.50 + 0.49 +0.85 +0.67 + 1.03 + 1.08 + 1.05 +0.64 +0.38 +0.98 +0.80 +oso
+2.70 +2.72 +2.62 +2.99 +2.76 +2.58 + 2.48 +2.49
+0.81 +0.13 + 0.68 + 0.62 + 0.62 +0.02 +0.22 +0.09
+0.02 +0.31 +0.22 +0.07 +0.06 -0.10 +0.07 -to.07 +0.12
+0.03 -0.09 -0.11 -0.12 +0.24 -0.08 +0.10 +0.09
0.00
+0.18 +0.15 +0.05 +0.05 +0.06 +0.20 -0.05 -0.36 -0.10 -0.13 +0.13 +0.13 +0.10 +0.04 -0.12 +0.13 -0.10 -0.15
+0.36 -0.32 -0.07 +0.07 +0.07 +0.05 +0.28 +0.16
+ 3.42
+3.19 +3.42 +3.28
-0.01 - 1.06
-0.13 -0.98 - 0.08 - 1.08 - 0.33 - 1.28
+3.38 +3.33
+0.02 -0.98 +0.14 -0.86
+2.82 +3.25
- 0.30 -1.20 +0.23 -0.67
+3.46 - 0.24 - 1.09
+3.21 +3.46 +3.22 +3.52
+0.51 -0.39 -0.61 - 1.26 -0.53 -0.93 -0.25 -0.70
+3.23 -0.70 -0.67 +3.51 -0.64 -0.58 +3.29 -0.69 -0.62
INTERGLACIAL OCEANIC 13C RECORD 231
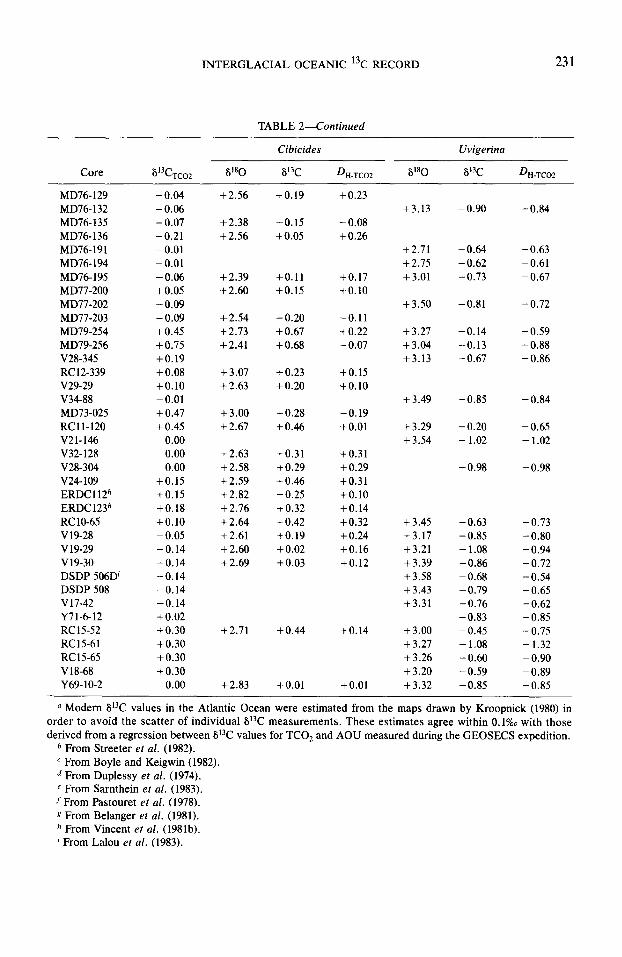
TABLE 2-Continued
Core V3C TC02 V80 -
MD76-129 -0.04 MD76-132 -0.06 MD76-135 -0.07 MD76-136 -0.21 MD76-191 -0.01 MD76- 194 -0.01 MD76-195 -0.06 MD77-200 +0.05 MD77-202 -0.09 MD77-203 -0.09 MD79-254 +0.45 MD79-256 + 0.75 V28-345 +0.19 RC12-339 +0.08 v29-29 +0.10 V34-88 -0.01 MD73-025 + 0.47 RCll-120 +0.45 V21-146 0.00 V32-128 0.00 V28-304 0.00 v24-109 +0.15 ERDCl 12h +0.15 ERDC123h +0.18 RC 10-6.5 +0.10 V19-28 -0.05 v19-29 -0.14 v19-30 -0.14 DSDP 506D’ -0.14 DSDP 508 -0.14 V17-42 -0.14 Y71-6-12 +0.02 RC15-52 +0.30 RC15-61 +0.30 RC15-65 +0.30 V18-68 + 0.30 Y69-10-2 0.00
+2.56 +0.23
+2.38 + 2.56
-0.15 +0.05
-0.08 +0.26
+ 2.39 +2.60
+0.11 +0.15
+0.17 +0.10
+ 2.54 +2.73 +2.41
-0.20 +0.67 +0.68
-0.11 +0.22 -0.07
+3.07 +2.63
+0.23 +0.20
+0.15 +0.10
+3.00 -t 2.67
+0.28 +0.46
-0.19 +0.01
+ 2.63 +0.31 fO.31 + 2.58 +0.29 +0.29 + 2.59 +0.46 +0.31 +2.82 +0.25 +0.10 +2.76 + 0.32 +0.14 +2.64 +0.42 +0.32 +2.61 +0.19 +0.24 + 2.60 +0.02 +0.16 + 2.69 +0.03 +0.12
+2.71
-t 2.83
+0.44
+0.01
+0.14
+0.01
- Cibicides Uvigerina
DH-TCO2 6'80 8'3C D~-~~~2 -
+3.13 -0.90 -0.84
+2.71 -0.64 -0.63 +2.75 -0.62 -0.61 +3.01 -0.73 -0.67
+3.50 -0.81 - 0.72
+3.27 -0.14 - 0.59 +3.04 -0.13 -0.88 +3.13 -0.67 -0.86
+3.49 -0.85 -0.84
+3.29 -0.20 - 0.65 +3.54 - 1.02 -1.02
-0.98 -0.98
+3.45 +3.17 +3.21 +3.39 +3.58 +3.43 +3.31
+3.00 +3.27 +3.26 +3.20 +3.32
-0.63 - 0.73 - 0.85 -0.80 - 1.08 -0.94 -0.86 -0.72 -0.68 -0.54 -0.79 -0.65 -0.76 -0.62 -0.83 - 0.85 -0.45 - 0.75 - 1.08 - 1.32 -0.60 -0.90 - 0.59 -0.89 -0.85 -0.85
u Modern 6i3C values in the Atlantic Ocean were estimated from the maps drawn by Kroopnick (1980) in order to avoid the scatter of individual Si3C measurements. These estimates agree within 0.1% with those derived from a regression between 8i3C values for TCO, and AOU measured during the GEOSECS expedition.
b From Streeter et al. (1982). c From Boyle and Keigwin (1982). d From Duplessy et al. (1974). e From Sarnthein et a/. (1983). f From Pastouret et al. (1978). g From Belanger et al. (1981). k From Vincent et al. (1981b). i From Lalou et al. (1983).

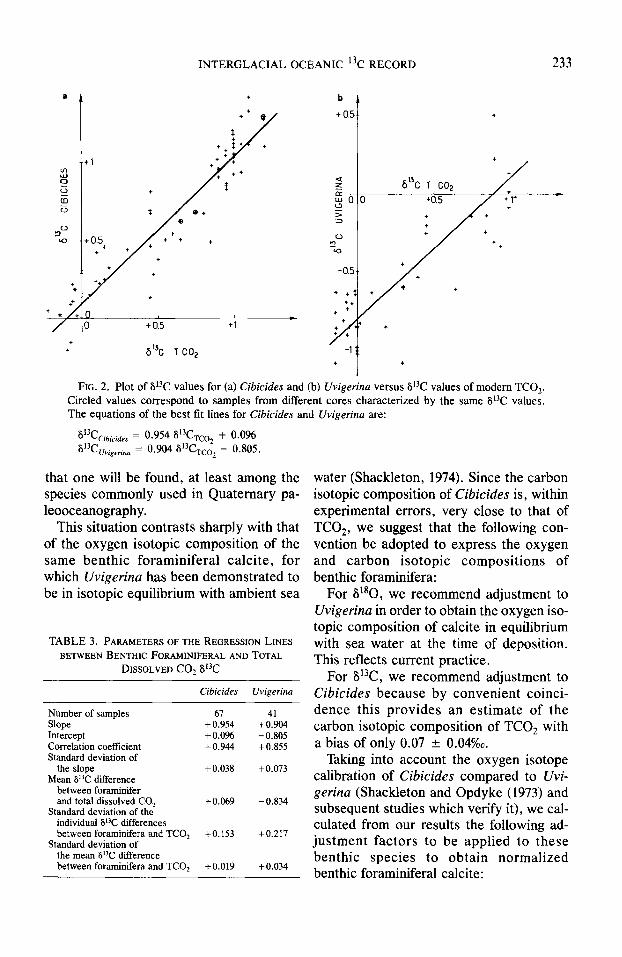
gerianus which have the same isotopic between benthic foraminiferal calcite and composition as C. wuellestorfi (we now TCO, would depend on the environmental shall refer to these only as Cibicides). Table conditions, we plotted S13C of recent Cibi- 2 also exhibits the calculated isotopic dif- tides (Fig. 2a) and Uvigerina (Fig. 2b) ferences between 613C of each benthic for- versus that of TCO,. The isotopic compo- aminiferal species and that of TCO*. We sition of benthic foraminifera and that of first calculated the mean value of these dif- TCO, are both subject to experimental er- ferences, their standard deviation (u), and rors. We therefore calculated the least- the standard deviation of the mean (c)); they squares solution for the regression lines fol- are lowing the procedure derived by York
for 67 Cibicides measurements: + 0.07 (1966). Results reported in Table 3 show
(u = 0.15; ?! = 0.02) that these regression lines are not statisti-
for 41 Uvigerina measurements: -0.83 cally different from one at the 66% conti-
(a = 0.22; cr = 0.034). dence level (1 a) and a fortiori at the 95% confidence level (2 u). We thus infer that
These results show that, at the 95% conti- Cibicides and Uvigerina are both good sen- dence level, 6i3C values of Cibicides differ sors of the carbon isotopic composition of from those of TCO, by only +0.07 +- TCO, over the whole range of its natural 0.04%0, in agreement with previous obser- variations. Cibicides is isotopically heavier vations of Belanger et al. (1981) and of than Uvigerina by 0.90 + 0.0%0 at the 95% Graham et al. (1981). The difference for confidence level and is more highly corre- Uvigerina (-0.83 + 0.07%0) is, however, lated with the bottom water conditions. The highly significant. 613C difference between Cibicides and Uvi-
The geographic distribution of Cibicides gerina calculated in that manner is in per- and Uvigerina is not uniform under modern fect agreement with comparisons indepen- conditions. In the Atlantic Ocean, below dently made by Shackleton and Cita (1979) 2000 m, Cibicides is frequent whereas Uvi- and by Blanc and Duplessy (1983) between gerina is rarely found. This latter species these species in many different sediments becomes an important component of the of varying age and original water tempera- fauna when the oxygen content of water ture. It therefore appears to be reliable for falls to below 5 ml/liter (Streeter and Shack- Tertiary and Quaternary specimens. leton, 1979). In fact, this species is very rare in the well-oxygenated core of NADW Isotopic Composition of Benthic
with high 613C values, but it is present in Foraminiferal Calcite and Bottom
Circumpolar Deep Water (CDW) and in Pa- Water Characteristics
cific Deep Water (PDW), which are rela- According to Emrich et al. (1970), 613C tively low in dissolved oxygen and 613C. values for CaCO, in isotopic equilibrium Therefore, the calibration of Uvigerina in are expected to be about 1.5%0 heavier than the range of high S13C values has been ob- those of dissolved bicarbonate. Our data, tained on only a few samples which might like those of the literature previously cited, correspond to individuals which have lived show that the carbon isotopic composition either under stressed conditions or in a of the foraminiferal calcite in Uvigerina and local microenvironment for which our 613C Cibicides is not in isotopic equilibrium with estimates are likely to be poor. The same that of TCO;?. At present, none of the other considerations also apply to Cibicides, analyzed benthic foraminiferal species has which is often rare in the low-oxygen, low- been reported to be in isotopic equilibrium 613C Pacific and Indian deep waters. with TCO, (Vincent et al., 1981a; Graham
In order to check if the 613C difference et al., 1981), and it seems highly unlikely
232 DUPLESSY ET AL.
INTERGLACIAL OCEANIC 13C RECORD 233
b
+ 0.5 t
+ 6°C T CO 2
FIG. 2. Plot of V3C values for (a) Cibicides and (b) Crvigerina versus 613C values of modem TCO,. Circled values correspond to samples from different cores characterized by the same 6t3C values. The equations of the best fit lines for Cibicides and Uvigerina are:
613C Crbrades . PC
= 0 954 8’3CTco, + 0.096 = 0.904 PC uv,gmna TC02 - 0.805.
that one will be found, at least among the species commonly used in Quaternary pa- leooceanography.
This situation contrasts sharply with that of the oxygen isotopic composition of the same benthic foraminiferal calcite, for which Uvigerina has been demonstrated to be in isotopic equilibrium with ambient sea
TABLE 3. PARAMETERSOFTHE REGRESSION LINES BETWEENBENTHK FORAMINIFERAL ANDTOTAL
DISSOLVED CO2 St3C
Cibicides Uvigerina
Number of samples 6-l Slope +0.954 Intercept +0.096 Correlation coefficient +0.944 Standard deviation of
the slope +0.038 Mean 8°C difference
between fornminifer and total dissolved CO2 +0.069
Standard deviation of the individual 6°C differences between foraminifera and TCO, +0.153
Standard deviation of the mean 6°C difference between foraminifera and TCO, +0.019
41 +0.904 -0.805 +0.855
+0.073
-0.834
+0.217
+0.034
water (Shackleton, 1974). Since the carbon isotopic composition of Cibicides is, within experimental errors, very close to that of TCO,, we suggest that the following con- vention be adopted to express the oxygen and carbon isotopic compositions of benthic foraminifera:
For 6180, we recommend adjustment to Uvigerina in order to obtain the oxygen iso- topic composition of calcite in equilibrium with sea water at the time of deposition. This reflects current practice.
For 613C, we recommend adjustment to Cibicides because by convenient coinci- dence this provides an estimate of the carbon isotopic composition of TCO, with a bias of only 0.07 + 0.04%0.
Taking into account the oxygen isotope calibration of Cibicides compared to Uvi- gerina (Shackleton and Opdyke (1973) and subsequent studies which verify it), we cal- culated from our results the following ad- justment factors to be applied to these benthic species to obtain normalized benthic foraminiferal calcite:

234 DUPLESSY ET AL.
Uvigerina: 0.00 for al80 + 0.90 for 613C Cibicides: + 0.64 for al80 0.00 for 613C.
All other living benthic species can be cal- ibrated following the method we developed in this study.
DISTRIBUTION OF BENTHIC 613C DURING THE LAST INTERGLACIATION
Since we demonstrated that the carbon isotopic composition of benthic foramini- fera reflects that of TCO,, in this section we now reconstruct the geographic distri- bution of the carbon isotopic composition of TCO, during the last interglaciation by measuring 6i3C of benthic foraminifera that lived in the major basins of the world ocean during isotopic substage 5e. We then use this reconstruction to deduce the major trend of the deep water circulation at that time and to compare it to the modern one.
Data Base
We used the cores in which benthic for- aminifera have been analyzed as a part of the study of the ocean during the last inter- glaciation (CLIMAP Project Members, 1984). Analytical values of all 613C anal- yses, relative to PDB, are reported in Ap- pendix 2 of the CLIMAP paper (CLIMAP Project Members, 1984).*
From all the available analyses, we se- lected only those of Uvigerina and of Cib- icides because we have not been able to calibrate the other species accurately. We had only a few analyses of recent speci- mens of Globocassidulina subglobosa and Nonion sp; the correlation between their S13C and that of TC02 was much too weak to permit any calibration. More analyses will be necessary to check the reliability of these two species. The carbon isotopic composition of Pyrgo in a deep-sea core is always much more scattered than that of the other accompanying species (CLIMAP
2 Readers may request Appendix 2, by written re- quest, from William E Ruddiman, Lamont-Doherty Geological Observatory of Columbia University, Pal- isades, N.Y. 10964.
Project Members, 1984, Appendix 2), and we do not recommend using this genus for isotopic work.
In Table 4, we report the raw 613C values of Uvigerina and Cibicides at the 5e level and the value of the benthic foraminiferal calcite (normalized to Cibicides by adding 0.90%0 to Uvigerina values). We increased our coverage by adding a few cores with good records of isotopic substage 5e which were not used for the CLIMAP reconstruc- tion of the last interglacial ocean (cores CH73-139C, DAL114, DSDP 508, MD73- 026, MD79-254, N079-28, SU81-32, V19- 30, V27-60, M13-519). Their locations are reported in Table 1 and the benthic fora- miniferal isotopic analyses in Table 4. In each core, the 5e level (reported in Table 4) was determined by selecting the lightest al80 value from among the benthic fora- miniferal analyses. Moreover we also checked that the #*O value at this level was typical of 5e, that is, similar to Holocene values ? 0.15%0, and rejected a few samples which did not pass this test (cores D-117, RCll-86, V22-38). In a few Atlantic cores with low sedimentation rates, some dis- agreement was observed between Uvi- gerina and Cibicides 6180 values, the first species being significantly heavier in 180. We believe that the occurrence of Uvi- gerina at these levels is due to bioturbation, because this species is generally absent in the Atlantic Ocean during the last intergla- ciation and we therefore took only into ac- count the Cibicides data.
Table 4 also displays two calculated dif- ferences:
he-TCOZ is the difference between 613C of benthic foraminiferal calcite during the last interglaciation and that of modern TCO,, estimated at the location and depth of the core.
D 5e-Holocene is the difference between 613C of benthic foraminiferal calcite and that of Holocene benthic calcite from the same core, or from a very nearby core raised from a similar depth.
The comparison of these two differences
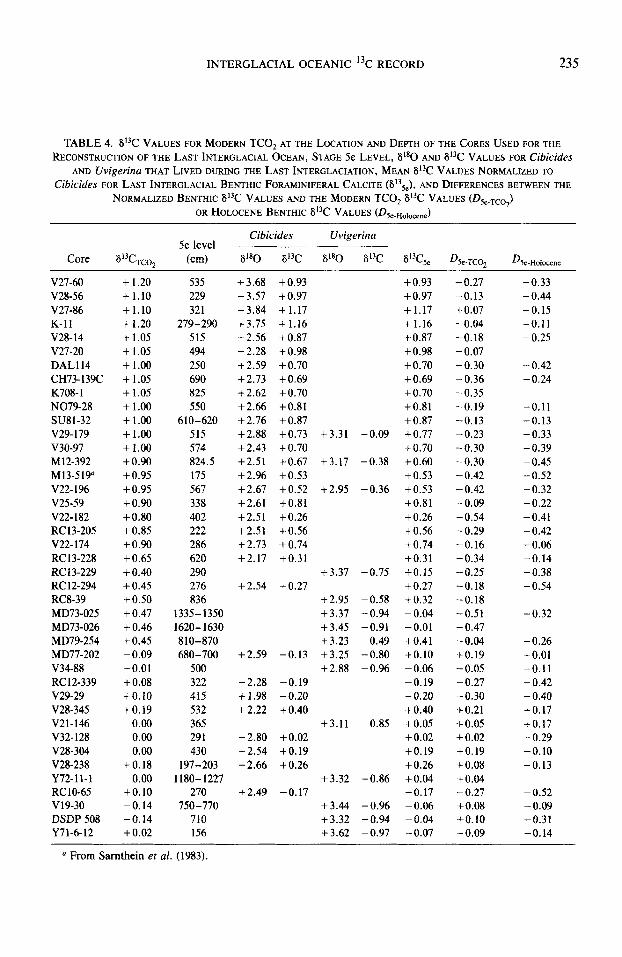
INTERGLACIAL OCEANIC 13C RECORD 235
TABLE 4. 613C VALUES FOR MODERN TCO, AT THE LIGATION AND DEFTH OF THE CORES USED FOR THE RECONSTRUCTION OF THE LAST INTERGLACIAL OCEAN, STAGE Se LEVEL, tY80 AND EiL3C VALUES FOR Cibicides
AND Uvigerina THAT LIVED DURING THE LAST INTERGLACIATION, MEAN G3C VALUES NORMALIZED TO Cibicides FOR LAST INTERGLACIAL BENTHIC FORAMINIFERAL CALCITE (S135e), AND DIFFERENCES BETWEEN THE
NORMALIZED BENTHIC 613C VALUES AND THE MODERN TCO, 613C VALUES (I),,.,,,) OR HOLOCENE BENTHIC 8°C VALUES (DJe.Ho,ocene)
Core 6i3C TC02
5e level (cm)
Cibicides Uvigerina
6’8O 613C 6’8O 6’)C iY3C 5e D5e-TC02 D Je-Holocene
V27-60 + 1.20 535 V28-56 + 1.10 229 V27-86 +1.10 321 K-11 + 1.20 279-290 V28-14 + 1.05 515 V27-20 + 1.05 494 DAL1 14 + 1.00 250 CH73-139C + 1.05 690 K708- 1 + 1.05 825 N079-28 +1.00 550 SUSl-32 +1.00 610-620 V29-179 +1.00 515 v30-97 + 1.00 574 Ml2-392 +0.90 824.5 Ml3-519” +0.95 175 V22-196 +0.95 567 V25-59 + 0.90 338 V22-182 +0.80 402 RCl3-205 +0.85 222 V22-174 +0.90 286 RCl3-228 +0.65 620 RCl3-229 +0.40 290 RC 12-294 + 0.45 276 RC8-39 +oso 836 MD73-025 +0.47 1335- 1350 MD73-026 + 0.46 1620-1630 MD79-254 +0.45 810-870 MD77-202 -0.09 680-700 V34-88 -0.01 500 RCl2-339 +0.08 322 V29-29 +0.10 415 V28-345 +0.19 532 V21-146 0.00 365 V32-128 0.00 291 V28-304 0.00 430 V28-238 +0.18 197-203 Y72-11-l 0.00 1180-1227 RClO-65 +0.10 270 v19-30 -0.14 750-770 DSDP 508 -0.14 710 Y71-6-12 +0.02 156
+3.68 +0.93 +3.57 +0.97 +3.84 +I.17 +3.7.5 + 1.16 +2.56 +0.87 +2.28 +0.98 + 2.59 +0.70 +2.73 +0.69 -!-2.62 +0.70 +2.66 +0.81 +2.76 +0.87 + 2.88 +0.73 +2.43 +0.70 +2.51 +0.67 +2.96 +0.53 i-2.67 -10.52 +2.61 +0.81 +2.51 +0.26 +2.51 +0.56 +2.73 +0.74 +2.17 +0.31
+2.54 +0.27
+2.59 -0.13
+3.31 - 0.09
+3.17 -0.38
+2.95 -0.36
i-3.37 -0.75
+2.95 -0.58 +3.37 - 0.94 +3.45 -0.91 +3.23 -0.49 +3.25 -0.80 +2.88 -0.96
+2.28 -0.19 + 1.98 -0.20 f2.22 +0.40
+3.11 -0.85 +2.80 +0.02 +2.54 +0.19 +2.66 + 0.26
+2.49 -0.17 +3.32 -0.86
+3.44 -0.96 + 3.32 - 0.94 +3.62 -0.97
+0.93 -0.27 +0.97 -0.13 +1.17 + 0.07 + 1.16 -0.04 +0.87 -0.18 + 0.98 -0.07 +0.70 -0.30 +0.69 -0.36 +0.70 -0.35 +0.81 -0.19 +0.87 -0.13 +0.77 -0.23 +0.70 -0.30 +0.60 -0.30 + 0.53 -0.42 +0.53 -0.42 +0.81 - 0.09 +0.26 -0.54 +0.56 -0.29 +0.74 -0.16 +0.31 - 0.34 +0.15 -0.25 fO.27 -0.18 +0.32 -0.18 -0.04 -0.51 -0.01 - 0.47 +0.41 -0.04 +0.10 +0.19 -0.06 -0.05 -0.19 -0.27 -0.20 -0.30 +0.40 +0.21 +0.05 +0.05 +0.02 +0.02 +0.19 +0.19 +0.26 +0.08 +0.04 + 0.04 -0.17 -0.27 -0.06 +0.08 -0.04 +0.10 -0.07 -0.09
-0.33 -0.44 -0.15 -0.11 - 0.25
-0.42 -0.24
-0.11 -0.13 -0.33 -0.39 -0.45 -0.52 - 0.32 - 0.22 -0.41 -0.42 - 0.06 -0.14 -0.38 - 0.54
-0.32
-0.26 +0.01 -0.11 -0.42 -0.40 +0.17 +0.17 -0.29 -0.10 -0.13
-0.52 -0.09 -0.31 -0.14
0 From Samthein et al. (1983).

236 DUPLESSY ET AL.
provides another check of the reliability of benthic foraminifena to record TC02 613C variations.
Discussion of the Mean Signal
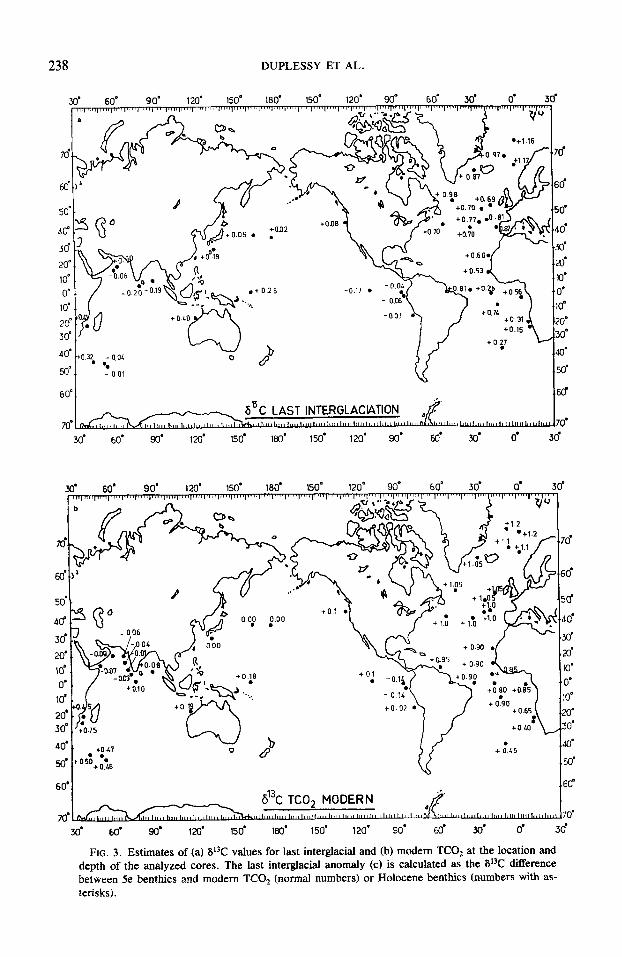
Figure 3a shows a map of the 613C values of benthic foraminiferal calcite as an esti- mate of the carbon isotopic composition of TCO, in the last interglacial ocean. This map is to be compared with a similar one showing the 613C values of modern TCO, at the same core location and depth (Fig. 3b), found in Tables 1 and 2. To make the comparison easier, Figure 3c shows the 613C difference between stage 5e benthics and either Holocene benthics (DSe~Holocene) or modern TCO, values (DSeFTCo2) or both.
These data show that all estimates of TCO, 613C in the Atlantic Ocean, in the Cir- cumpolar Deep Water, and in the Norwegian Sea (with the exception of one value) are significantly lower (isotopically lighter) than those of today. This conclusion holds when the stage 5e benthic foraminiferal cal- cite is compared both to the actual TCO, modern measurements and to Holocene benthic foraminiferal calcite. In the Pacific and Indian oceans, the 613C departure from modern conditions is much less important. We calculated the mean 613C difference be- tween the last interglaciation and today by averaging the values of DSesTC02 and D,,- Holocene in each major oceanic basin (Table 9. The DSemTCo2 values are significantly more positive ( + 0.14 + 0.10) than those of D Se-Holocene. This difference is due partly to experimental errors, partly to errors in the estimates of TCO, S13C, and partly to our choice of the 613C value of Cibicides as an estimate of the carbon isotopic composition of past TCO,, because we have previously shown that this genus is +0.07 + 0.04%0 isotopically heavier than TCO,. We there- fore took the mean value of DSemTCo2 and D 5e-Holocene as the best estimate of the 613C departure from modern conditions in each oceanic basin during the last interglacia- tion. The 613C difference between the last interglaciation and today is about -0.2%0 in
the Norwegian Sea and in the western At- lantic, whereas it is only -0.1%0 in the In- dian and Pacific oceans. The same differ- ences for the eastern Atlantic Ocean and for the Circumpolar Deep Water of the Southern Ocean are much larger (about -0.30%0). We shall discuss separately the global 613C decrease of the oceanic TC02 and the re- gional pattern.
Although our coverage is not uniform over the whole ocean, the data in Table 5 show that the bulk of the CO2 dissolved in the ocean had a 613C about 0.15 t 0. 12%0 lower than that of the modern one at the 95% confidence level (taking into account that the Pacific Ocean represents more than 50% of the world ocean water mass). Such a 613C decrease for the deep TCO, cannot be explained by enhanced production of particulate organic carbon in surface wa- ters, later oxidized at depth by bacteria, without any change in the biosphere: since surface waters represent less than 5% of the oceanic water mass, a 0.15 + 0.12%0 V3C decrease in deep waters would be balanced by a 3.0 ? 2.4%0 613C increase in surface waters. Such heavy values for surface water 613C have never been recorded by any planktonic foraminiferal isotopic record (Duplessy, 1982; CLIMAP Project Members, 1984, Appendix 2). Thus the oceanic impoverishment in 13C can only be explained by a depletion, compared to its present value of the global organic carbon reservoir, which is highly depleted in 13C.
Two mechanisms may explain this change: a reduction in the amount of or- ganic-rich sediment deposited on the con- tinental shelves or a decrease of the conti- nental biomass. Following Broecker (1982), we first test the assumption that the change in the isotopic composition of oceanic carbon resulted from a variation of the de- position of shelf sediment. The lower oceanic S13C would then be explained by a lesser deposition of organic-rich sediment on shelves and estuaries during the sea- level rise at the beginning of stage 5e than that observed during the Flandrian trans-

INTERGLACIAL OCEANIC 13C RECORD 237
gression at the beginning of the Holocene. In that case, less organic matter would have been trapped on the shelves, resulting in a higher TCO, and lower i3C content in the last interglacial ocean than in the modern one. We have no way to check this hypoth- esis directly since most oceanic shelves and estuaries were emptied during the last gla- ciation. However, the 125,000 yr B.P. sea level has been consistently reported to have been about 6 m above the present one (Bloom et al., 1974; Ku et al., 1974), re- sulting in a well-marked increase of shelf area along the Atlantic coast of the United States and along the margin of the Baltic Sea (Flint, 1971). Thus, no field evidence supports the hypothesis that shelves and estuaries were less efficient traps for or- ganic-rich sediments during the last inter- glaciation than during Holocene times.
A mean oceanic G3C value lower than today by 0.15 + 0.12%0 may also be ex- plained by a 7 + 6% reduction of the con- tinental biomass, in which the carbon is se- questered in forest and soil (Shackleton, 1977). An exact analysis of the history of the last interglacial vegetation is at present impossible since precise dating of the con- tinental deposits is not yet available. Al- though we are unable to document the amount of the Eemian-Sangamon biomass, an estimate lower than the modern one by a value as large as 7 + 6% would have to be explained in terms of the tropical vege- tation rather than the temperate regions. We find this the most plausible way to ex- plain the change in carbon isotopic com- position of the CO, dissolved in the ocean during the last interglaciation
Regional Variations
We turn now to the regional differences in 6r3C between the various oceanic basins. One of the most striking differences be- tween the last interglacial and the modern patterns is the low 613C values found in the eastern Atlantic Ocean. The present hy- drography of this basin provides some clues
to the origin of this pattern. The deep water circulation in the eastern Atlantic Ocean is distinctly different from and much weaker than that of the western Atlantic (Wor- thington, 1976). The geochemical proper- ties of the water below 1500 m in the eastern basin depend on a balance between the inflow of NADW entering from the north and of Antarctic Bottom Water (AABW), entering from the western basin through the Romanche and Vema fracture zones in the equatorial latitudes. From there, AABW flows north and south and mixes with NADW. The relatively low input of particulate organic matter from surface water in the Atlantic Ocean results in weak oxygen utilization within the body of NADW; consequently the 613C pattern of TCO, mainly reflects the physical mixing between NADW and AABW (Kroopnick, 1980). No geological evidence exists for any drastic difference in oceanic produc- tivity between the last interglaciation and the modern ocean. Therefore, the low 613C values observed in the eastern Atlantic Ocean during stage 5e imply that the influ- ence of AABW was much stronger than that of NADW.
Such a pattern contrasts with the present one, since the modern average production rates of AABW and NADW are generally assumed similar (Kuo and Veronis, 1973). The enhanced intluence of AABW during the last inter-glaciation should also be ob- served in the southern basin of the western Atlantic Ocean. However, we have no core from this area with a high sedimentation rate and reliable benthic isotopic record (Fig. 3a).
We shall now use the results of numerical calculations made with the model of Kuo and Veronis (1970, 1973) to speculate about the causes of the east-west S13C differ- ences measured in the last interglacial ocean and of the lower 613C values found in the Southern Ocean during stage 5e. The increased relative influence of AABW in the eastern Atlantic Ocean during the last interglaciation can be explained by either
238 DUPLESSY ET AL.
50. 60' 90" 120' 150' lfld 150' 120' 90' bd 30’ 0’ 3d
\ a /3 A r-Y.- GiGi-lL
I uu
B
v
+om . . -v-u: k
10' 2d 7 O. 0
40* I ~--“~‘s~~~.:~.,,~+026 -0.1, - o.c6 .
. l OLO
0 2
10" -0.01
l v + 0.7L +0.3l 20’
3d +0.15
30" + 0
‘. 032 l -0OL 0 50” ‘9
- 0.01
v 40'
50’
60'
70.
30' b0’ 90’
6d
70'
120’ 150’ Id 150' 120' 90' 60' 30' 0' 3d
30' 60' 90' 60’ 30’
60' 60"
70. 70.
30' bo' 90' 120' 150' 180' 150' 120' 90' Ed 30' 0' 3d
FIG. 3. Estimates of (a) 613C values for last interglacial and (b) modem TCO, at the location and depth of the analyzed cores. The last interglacial anomaly (c) is calculated as the 6°C difference between 5e benthics and modem TCO, (normal numbers) or Holocene benthics (numbers with as- terisks).

INTERGLACIAL OCEANIC 13C RECORD 239
FIG. 3-Continued
enhanced AABW production or reduced NADW production. To resolve this ambi- guity, we note that the west-east 613C gra- dient was stronger than the modern one during the last interglaciation, with high 6t3C in the west and low 613C in the east (compare Figs. 3a and b). This gradient re- flects the role of western boundary currents in the deep water dynamics. Since the large-scale features of the 6°C variations in the whole ocean closely follow the distri- bution in dissolved oxygen (Kroopnick, 1974, 1980), we can interpret the last inter- glacial 613C distribution within the frame- work of numerical experiments by Kuo and Veronis (1973) and use the global distribu- tion of dissolved oxygen as a tracer of ad- vection and diffusion processes in the abyssal ocean. In this model, which does not take into account topographic effects, the oxygen distribution is shown to depend on only three independent parameters: (a) the consumption rate of dissolved oxygen in abyssal waters (since we have no way to estimate this in the past, we shall assume it
is constant); (b) the Peclet number, propor- tional to the ratio of the magnitude of the upwelling velocity, assumed uniform, to the eddy diffusion (this parameter is a measure of advection to diffusion); (c) the recircu- lation, the flux which measures the east- ward transport in the antarctic circumpolar current.
The Kuo and Veronis numerical calcu- lations combine the advective-diffusive equation with Stommel’s (1958) abyssal cir- culation model for an idealized world ocean basin. They show that a significant rotation of the oxygen contours toward a north- south orientation, resulting in an increase of the west-east oxygen gradient, reflects only an increase of the Peclet number and thus of the advection because the major water transport occurs along the western margin of the oceanic basins. Therefore these model data imply that the 613C distri- bution during the last interglaciation re- flects an enhanced AABW production more than a reduced NADW production.
The same circulation model can also be

240 DUPLESSY ET AL.
TABLE 5. REGIONAL 613C DIFFERENCES BETWEEN LAST INTERGLACIATION AND TODAYS
D5e-TCOZ D Se-Holocene 5e anomaly (U;is) b;3 (4
Norwegian Sea -0.03 (0.18; 0.09) -0.26 (0.15; 0.08) -0.15 (0.08) Western Atlantic -0.16 (0.11; 0.06) -0.29 (0.11; 0.06) -0.16 (0.06) Eastern Atlantic -0.32 (0.16; 0.04) -0.30 (0.14; 0.04) -0.31 (0.04) Indian Ocean -0.04 (0.22; 0.09) -0.16 (0.23; 0.10) -0.10 (0.09) Circumpolar water -0.34 (0.18; 0.09) -0.34 (0.09) Pacific Ocean +0.03 (0.13; 0.03) - 0.18 (0.20; 0.07) -0.08 (0.05) World Ocean -0.15 (0.06)
a &-TCO~ is the 8°C difference between 5e benthic foraminiferal calcite and modem TCO,. D5e.Ho,ofene is the 8i3C difference between 5e and Holocene benthic foraminiferal calcite. (T is the standard deviation of each individual value and 0 is the standard deviation of the mean value. The 5e anomaly is calculated as the mean value between the two differences.
used to interpret the low 6i3C values found in last interglacial CDW benthic foramini- fera. In the circumpolar current, mixing oc- curs between deep water rising from all of the southern oceans and surface water with a high dissolved oxygen concentration and a high 613C value. The deep water is low in dissolved oxygen and low in 613C due to extensive nutrient regeneration and oxida- tion of organic matter. The occurrence of a significant 613C decrease in the last inter- glacial CDW compared to the present one might thus be explained by increased oxi- dation of organic matter reflecting en- hanced local productivity in the surface wa- ters. However, the sedimentation rates in subantarctic cores MD73-025 and MD73- 026 were lower during isotope stage 5e than during the Holocene, which suggest that the local diatom productivity was also lower. The low 613C values measured in the last interglacial CDW benthic foraminifera are thus better explained by a change in the deep water circulation. Numerical calcula- tions with the model of Kuo and Veronis (1973) show that a decrease of eastward transport has the principal effect of de- creasing the oxygen concentration (and thus the Fi3C) in the circumpolar region be- cause of the longer residence time of the deep water mass. The model also shows that the lower oxygen concentration is as- sociated with a decrease in the eastern side of the Pacific Ocean. However, we do not yet have enough data in the Pacific Ocean
to measure accurately a small change in the west-east 613C gradient. It would be nec- essary to have a larger core coverage, mainly in the Pacific and Southern oceans, to compute the actual decrease in eastward transport in the last interglacial ocean.
CONCLUSION
Our results show that the 13C records of two benthic foraminifera, Uvigerina and Cibicides, provide a tool to estimate the carbon isotopic composition of TCO, in the past ocean. The direct comparison between modem TC02 613C and that of upper Ho- locene benthic foraminifera appears to be the most accurate way to check the reli- ability of benthic foraminifera as recorders of the carbon isotopic composition of TCO,. This method should be applied to other species as well as those we have an- alyzed in order to determine both their re- liability and the correction factor to apply to raw 613C measurements for obtaining es- timates of TCO, 6i3C.
The 13C distribution in the deep ocean during the last interglaciation differed no- ticeably from that at present. This shows how sensitive both the global carbon cycle and the deep water circulation are to minor changes in the earth’s climate. Although the last interglacial ocean was, in general, very similar in surface temperature to to- day’s ocean, the global TCO, had a lower 813C than the present one, reflecting prob- ably lesser development of the continental

INTERGLACIAL OCEANIC 13C RECORD 241
biosphere following the very severe Saa- lian-Illinoian glacial age. The deep water circulation was characterized by increased AABW production and enhanced north- south advection, resulting in an increase in the west-east 613C gradient in the Atlantic Ocean. By contrast, the eastward transport is likely to have been reduced. The differ- ences between the last interglaciation and the modern deep water circulations are per- haps more important than those determined for the surface realm of the ocean.
ACKNOWLEDGMENTS
Thanks are due to J. Labeyrie and M. Bender for useful1 discussions and comments on the manuscript, to W. S. Broecker for a careful review of the manu- script, to R. H. Fillon for providing samples of core HU7558, and to J. Antignac, H. Leclaire, M. Hall, and B. Le Coat for their help in the isotopic analyses. The coring cruises of the French M/S Marion Dufresne were supported by TAAF, those of the French RIV Noroit, Suroit, and Jean Charcot by CNEXO. J. C. Duplessy was supported by CNRS-CEA and EEC Grant CLI-005F. N. J. Shackleton was supported by NERC Contract GR3-3606. Office of Naval Research Grant ONR N00014-75CO210 and NSF Grant OCE 78 25448 support the core collection of Lamont Doherty Geological Observatory.
REFERENCES
Belanger, P. E., Curry, W. B., and Matthews, R. K. (1981). Core-top evaluation of benthic foraminiferal isotopic ratios for paleo-oceanographic interpreta- tions. Palaeogeography, Palaeoclimatology, Pa- laeoecology 33, 205-220.
Blanc, P. L. (1981). “Paleoclimatologie isotopique et histoire de l’eau profonde atlantique depuis 15 mil- lions d’annees.” Thesis, University of Paris XI.
Blanc, P. L., and Duplessy, J. C. (1983). The deep water circulation during the Neogene and the impact of the Messinian salinity crisis. Deep Sea Research 29, 1391-1414.
Bloom, A. L., Broecker, W. S., Chappell, J. M. A., Matthews, R. K., and Mesolella, J. (1974). Quater- nary sea level fluctuations on a tectonic coast: New 230Th/234U dates from the Huon Peninsula, New Guinea. Quaternary Research, 4, 185-205.
Boersma, A., and Shackleton, N. (1977). Tertiary ox- ygen and carbon isotope stratigraphy, site 357 (mid latitude south Atlantic). In “Initial Reports of the Deep Sea Drilling Project.” Vol. 39, pp. 911-924. U.S. Govt. Printing Office, Washington, D.C.
Broecker, W. S. (1982). Glacial to interglacial changes
in ocean chemistry. In “Progress in Oceanography” (E. Kraus, Ed.), Vol. 11, pp. 151-197.
CLIMAP Project Members (1984). The last interglacial ocean. Quaternary Research 21, 123-224.
Craig, H. (1970). Abyssal carbon-13 in the South Pa- cific. Journal of Geophysical Research 75,691-695.
Curry, W. B., and Lohmann, G. P. (1982). Carbon iso- topic changes in benthic foraminifera from the western South Atlantic: Reconstruction of glacial abyssal circulation patterns. Quaternary Research, 18, 218-235.
Duplessy, J. C. (1972). “La geochimie des isotopes stables du carbone dans la mer.” Thesis, University of Paris VI.
Duplessy, J. C. (1978). Isotope Studies. In “Climatic Change” (J. Gribbin Ed.), pp. 46-67. Cambridge Univ. Press, London/New York.
Duplessy, J. C. (1982) Circulation des eaux profondes Nord Atlantiques au tours du demier cycle clima- tique, Bulletin de Znstitut de Geologie du Bassin, d’dquitaine 31, 379-391.
Duplessy, J. C., Chenouard, L., and Reyss, J. L. (1974). Paltotemperatures isotopiques de l’Atlan- tique Equatorial. In “Colloque International du CNRS N”219,” pp. 251-258. CNRS, Paris.
Duplessy, J. C., Chenouard, L., and Vila, F. (1975). Weyl’s theory of glaciation supported by isotopic study of Norwegian core K-11. Science 188, 1208- 1209.
Duplessy, J. C., Lalou, C., and Vinot, A. C. (1970). Differential isotopic fractionation in benthic fora- minifera and paleotemperatures reassessed. Science 168, 250-251.
Duplessy, J. C., Moyes, J., and Pujol, C. (1980). Deep water formation in the North Atlantic Ocean during the last ice age. Nature (London), 286, 479-482.
Emrich, K., Ehhalt, D. H., and Vogel, J. C. (1970). Carbon isotope fractionation during the precipita- tion of calcium carbonate. Earth and Planetary Sci- ence Letters 8, 363-370.
Flint, R. F. (1971). “Glacial and Quaternary Ge- ology.” Wiley, New York.
Fontugne, M. F., and Duplessy, J. C. (1981). Organic carbon isotopic fractionation by marine plankton in the temperature range - 1 to 31°C. Oceanologica Acta 4, 85-90.
Graham, D. W., Corliss, B. H., Bender, M. L., and Keigwin, L. D. (1981). Carbon and oxygen isotopic disequilibrium of recent deep sea benthic foramini- fera. Marine Micropaleontology 6, 483-497.
Hutson, W. H. (1980) Bioturbation of deep sea sedi- ments: Oxygen isotopes and stratigraphy uncer- tainty. Geology 8, 127-130.
Kellogg, T. B. (1976). Late Quaternary climatic changes: Evidence from deep sea cores of Norwe- gian and Greenland Seas. In “Investigation of Late Quaternary Paleooceanography and Paleoclima- tology” (R. M. Cline and J. D. Hays, Eds.), Geo- logical Society of America Memoir, 145, 77- 110.

242 DUPLESSY ET AL.
Kellogg, T. B., Duplessy, J. C., and Shackleton, N. J. (1978). Planktonic foraminiferal and oxygen isotopic stratigraphy and paleoclimatology of Norwegian Sea deep-sea cores. Boreas 7, 61-73.
Kroopnick, P. (1974). The dissolved O,-CO,-t3C system in the eastern equatorial Pacific. Deep Sea Research 21, 211-222.
Kroopnick, P. (1980). The distribution of t3C in the Atlantic ocean. Earth and Planetary Science Letters 49, 469-484.
Kroopnick, P., Deuser, W. G., and Craig, H. (1970). Carbon- 13 measurements on dissolved inorganic carbon at the North Pacific (1969) Geosecs Station. Journal Geophysical Research 75, 7668-7671.
Kroopnick, P., Weiss, R. E and Craig, H. (1972). Total CO,, t3C and dissolved ‘*O at Geosecs II in the North Atlantic. Earth and Planetary Science Letters 16, 103-l 10.
Ku, T. L., Kimmel, M. A., Easton, W. H., and O’Neil, T. J. (1974). Eustatic sea level 120,000 years ago on Ohau, Hawaii. Science 183, 959-962.
Kuo, H. H., and Veronis, G. (1970). Distribution of tracers in the deep oceans of the world. Deep Sea Research 17, 29-46.
Kuo, H. H., and Veronis, G. (1973). The use of ox- ygen as a test for an abyssal circulation model. Deep Sea Research 20, 871-888.
Lalou, C., Brichet, E., Leclaire, H., and Duplessy, J. C. (1983). Uranium series disequilibrium and iso- tope stratigraphy in hydrothermal mounds samples from D.S.D.P. sites 506-509, leg 70, and site 424, leg 54: An attempt at chronology. In “Initial Reports of the Deep Sea Drilling Project,” Vol. 70. U.S. Govt. Printing Offtce, Washington, D.C.
Lohmann, G. P. (1978) Response of the deep sea to ice ages. Oceanus 21(4), 58-64.
Pastouret, L., Charnley, H., Delibrias, G., Duplessy, J. C., and Thiede, J. (1978). Late Quatemary cli- matic changes in western tropical Africa deduced from deep-sea sedimentation off the Niger delta. Oceanologica Acta 1, 217-232.
Peng, T. H., Broecker, W. S., Kipphut, G., and Shackleton, N. J. (1977). Benthic mixing in deep sea cores as determined by t4C dating and its implica- tions regarding climate, stratigraphy and the fate of fossil fuel CO,. In “The Fate of Fossil Fuel CO, in the Oceans” (N. R. Anderson and A. Malahoff, Eds.), pp. 355-373. Plenum, New York.
Richard, F. A. (1965). Dissolved gases other than carbon dioxide. In “Chemical Oceanography” (J. P. Riley and G. Skirrow, Eds.), Vol. 1, pp. 197-225. Academic Press, New York.
Sackett, W. M., Eckelmann, W. R., Bender, M. L., and Be, A. W. H. (1965). Temperature dependence of carbon isotope composition in marine plankton and sediments. Science 148, 235-237.
Samthein, M., Erlenkeuser, H., von Grafenstein, R., and Schroder, C. (1983). Stable isotope stratigraphy
for the last 750,000 years: Meteor core 13-519 from the eastern equatorial Atlantic. “Meteor” For- schungsergebnisse, Reihe C, in press.
Savin, S. M., Douglas, R. G., Keller, G., Killingley, J. S., Shaughnessy, L., Sommer, M. A., Vincent, E., and Woodruff, F. (1981). Miocene benthic fora- miniferal isotope records: A synthesis. Marine Mi- cropaleontology 6, 423-450.
Schnitker, D. (1974). West Atlantic abyssal circulation during the past 120,000 years. Nature (London) 248, 385-387.
Schnitker, D. (1980). Quatemary deep-sea benthic for- aminifers and bottom water masses. Annual Review of Earth and Planetary Sciences 8, 343-370.
Shackleton, N. J. (1974). Attainment of isotopic equi- librium between ocean water and the benthonic for- aminifera genus Uvigerina: Isotopic changes in the ocean during the last glacial. In “Colioque Inter- national du CNRS N” 219,” pp. 203-209. CNRS, P&S.
Shackleton, N. J. (1977). Tropical rain forest history and the equatorial Pacific carbonate dissolution cy- cles. In “The Fate of Fossil Fuel CO, in the Oceans” (N. R. Anderson and A. Malahoff, Eds.) pp. 401-428. Plenum, New York.
Shackleton, N. J., and Cita, M. B. (1979). Oxygen and carbon isotope stratigraphy of benthic foraminifers at site 397: Detailed history of climatic change during the late Neogene. In “Initial Reports of the Deep Sea Drilling Project,” Vol. 47, pp. 433-444. U.S. Govt. Printing Office, Washington, D.C.
Shackleton, N. J., Imbrie, J., and Hall, M. (1983). Ox- ygen and carbon isotope record of east Pacific core V19-30: Implications for the formation of deep water in the Late Pleistocene North Atlantic. Earth and Planetary Science Letters, 65, 233-266.
Shackleton, N. J., and Opdyke, N. D. (1973). Oxygen isotope and paleomagnetic stratigraphy of equatorial Pacific core V28-238: Oxygen isotope temperatures and ice-volume on a 105-year scale. Quaternary Re- search 3, 39-55.
Shackleton, N. J., and Opdyke, N. D. (1976). Oxygen isotope and paleomagnetic stratigraphy of Pacific core V28-239 late Pliocene to latest Pliocene. In “Investigation of Late Quaternary Paleooceanog- raphy and Paleoclimatology” (R. M. Cline and J. D. Hays, Eds.), Geological Society of America, Memoir 145, 449-464.
Stommel, H. (1958). The abyssal circulation. Deep Sea Research 5, 80-82.
Streeter, S. S. (1973). Bottom water and benthonic foraminifera in the North Atlantic-Glacial-Inter- glacial contrasts. Quaternary Research 3, 131-141.
Streeter, S. S., Belanger, P. E., Kellogg, T. B., and Duplessy, J. C. (1982). Late Pleistocene Paleo- oceanography of the Norwegian-Greenland Sea: Benthic foraminiferal evidence. Quaternary Re- search 18, 72-90.

INTERGLACIAL OCEANIC 13C RECORD 243
Streeter, S. S., and Shackleton, N. J. (1979). Paleocir- culation of the deep North Atlantic: 150,000 year record of benthic foraminifera and oxygen-18. Sci- ence 203, 168-171.
Vincent, E., Killingley, J. S., and Berger, W. H. (1981a) Stable isotope composition of benthic fora- minifera from the equatorial Pacific. Nature (London) 289, 639-643.
Vincent, E., Killingley, J. S., and Berger, W. H. (198 1 b). Stable isotopes in benthic foraminifera from Ontong-Java plateau, box cores ERDC 112 and 123.
Palaeogeography, Palaeoclimatology, Palaeo- ecology 33, 221-230.
Woodruff, F., Savin, S. S., and Douglas, R. G. (1980). Biological fractionation of oxygen and carbon iso- topes by recent benthic foraminifera. Marine Micro- paleontology 5, 3- 11.
Worthington, L. V. (1976). “On the North Atlantic Circulation.” John Hopkins Press, Baltimore/ London.
York, D. (1%6). Least-square fitting of a straight line. Canadian Journal of Physics 44, 1079-1083.





























