Embed Size (px)
Citation preview

_< . .. i
MATRICULA:
TELEFONO :
JIICENCIATURA:
TRIMESTRE LECTIVO:
HRS. A LA SEMANA:
/TITULO:
INGENIERIA DE LOS FILIMENTOS
92-P
20 HORAS
AISLAMIENTO Y PURIFICACION DE ACTINIDINA DE Ac t i n i d i a c h i n e n s i s
M. EN O. SALVADOR R. TELLO SOLIS
LUGAR DE REALIZACION: LABORATORIO DE BIOFISICOQUIMICA DEPARTAMENTO DE QUIMICA. DIVISION DE CIENCIAS BASICAS E INGENIERIA.
FECHA DE INICIO:
/FECHA DE TERM I NAC ION I 1 I
CLAVE :
NOMBRE DEL PROYECTO: 1
8 DE JULIO DE 1991
7 DE FEBRERO DE 1 9 3
AISLAMIENTO Y PURIFICACEON DE FICTINIDINA DE A c t i n i d i a c h i n e n s i s
1. EN Q. SALVADOR R. TELLO SOLIS DEPARTAMENTO nE QUIMICA.

Casa abierta al tiempo
UNIVERSIDAD AUTONOMA METROPOLITANA Julio 3, 1992.
BIOL. MARCO AURELIO PEREZ HERNANDEZ. SECRETARIO ACADMICO DIVISION DE CIENCIAS BIOLOGICAS Y DE LA SALUD. P r e s e n t o .
Por medio de la presente informo a Usted que la alumna MARIA CRISTINA ALVAREZ MALFAVON, matrícula 87344730, de la Licenciatura en ingeniería de los alimentos terminó satisfactoriamente su proyecto de servicio social, bajo mi dirección.
La alumna ALVAREZ MALFAVON desempeñó su trabajo de servicio social con gran interés y dedicación, alcanzando cubrir todos los objetivos que se plantearon . Cabe señalar, que además fue co-autora de un trabajo de investigación relacionado directamente con su proyecto de servicio social y que se presentó en el VI Simposio de estudiantes de posgrado en Química "FERNANDO ROMO", celebrado los dias 28 y 29 de Mayo de 1992 en la Universidad de Guanajuato.
Sin más por el momento, le saluda
ATENTAMENTE. "CASA ABIERTA AL. TIEMPO"
I
M. eri Q. SALVADOR R. ELLO SOLIS. DEP TAMENTO DE QUIMICA. A S ?R E S O R.
UNIDAD IZTAPALAPA Av. Michoacán y La Purisima. Col. Vicentina. Irtapalapa. D.F. C.P. 09340. Tel.: 686-03-22 TELEFAX (5) 686-89-99 TELEX: UAMME 176496

I I I I I I I I I I I I II il
f
:
AISLAMIENTO Y PURIFICACION DE ACTINIDINA
DE Actinidia chinensis

AISLAMIENTO Y PURIFICACION DE ACTINIDINA . DE Actinidia chinensis
OBJETIVO GENERAL
Utilizaci6n de las técnicas de Cromatografia de Filtracibn en gel y Cromatografía líquida de alta presi6n (HPLC), en la purificacibn de proteínas.
OBJETIVOS PARTICULARES
X 4islamiento de la proteasa sulfhidrílica del fruto del Kiwi,Actinidia chinensis
X Purificación de la proteasa sulfhidrilica del fruto del Kiwi,Actinidia chinensis
rC Determinacibn de la actividad especifica.
JUSTIFICACION
Hoy en 4 1 es de suma np in , cono er can detalle la estructura y estabilidad de una proteína, tanto desde un punto de vista fundamental, como por sus aplicaciones biotecnolbgicas (alimentos y fdrmacos). Para realizar dichos estudios , es indispensable que la proteína se pueda aislar lo mas pura y homogeneamente posible.
En el presente proyecto de Servicio Social se colaboró en el aislamiento y la purificaci6n de una proteína (enzima) del fruto del Kiwi, Actinidia chinensis. La purificaci6n de esta enzima es esencial para poder continuar con estudios estructurales por espectroscopia y dicroismo circular.
.

INTRODUCCION
t Las proteasas sulfhidrilicas comprenden a un grupo de .' enzimas, en las cuales su actividad proteolítica depende de un grupo sulfhidrilo de un residuo de cistelna en su sitio activo. Este tipo de enzimas se encuentra ampliamente distribuido en diferentes especies de animales, vegetales y bacterias. Hay evidencia de que la mayoria de ellas pertenecen a una familia d e enzimas que presentan homologia, tanto estructural como funcional.(l)
Las proteasas sulfhidrílicas mds conocidas son las de origen vegetal, ya que dichas fuentes se obtienen con relativa facilidad Y en cantidades considerables. Este tipo de proteasas es ampliamente utilizado en la industria, por ejemplo en ablandamiento de carnes y clarificación de cerveza, su funci6n metabólica sugiere que en los vegetales actuan como agentes proteolíticos contra infecciones fiingicas. ( 2 )
Estudios sobre su reactividad indican que el sistema cisteína - histidina, en el sitio activo, parece, ser una r g r * i r 4 f r < _ _ - - . - - - . - - - s c i -a Clrni j " en las enzimas sulfhidrilicas de vegeta les . ( Z - < - r l .
I

, La papaina, proteasa sulfhidrilica de la papaya
tropica1,Carica DaDaya, es considerada como un ejemplo tipico de este tipo de proteasas vegetales, la molecula de esta proteina consiste de una sola cadena polipeptidica de 212 residuos de aminoácidos y tiene una masa molecular de 23 350 Da. La papaina es una proteasa sulfhidrílica a l a cual se le ha determinado su secuencia de aminoacidos y su estructura tridimensional por rayos x ( 7 , 8 ) .
Existe una gran variedad de proteasas sulfhidrílicas caracterizadas por sus masas moleculares y por sus puntos isoeléctricos, casi todas estas enzimas tienen masas moleculares entre 20 O00 y 33 O00 Da. Sus puntos isoeléctricos son muy variables lo que probablemente se debe a 105 diferentes medios biol6gicos en los cuales actuan.

METODOLOQICI
Diagrama di-? f h j r para .la p i~ i . i . t . i : . i u r i de l a e r i : z i m a I4 c t i n i d i a c h i n erzck?.
AISLAMIENTO
I,.- ........... 1
1 . . . .......... _..-._I_-.

DI AL I C 1 S
CENTRIFUGACION (En frío)
.
SOBHENADANTE (Almacenamiento a O O C )
4
I EXiHACTO CRUDO 1

PURIFICACION.
<
FILTRACION EXTRACTO CRUDO ( A través de membrana millipore)
EVALUACION DE Lñ
(Election de un -------l sustrato adecuado)
ACTIVIDAD ENZIMATICA.
.
EVALUACION DE LA ACTIVIDAD ENZIMATICA (Variacih activadora
y pH sustrato)
CROMATOGRAFIA DE FILTRACION EN GEL (CFG)
ELECCION DE FRACCIONES

REPRECIPITACION w DI AL IC1 C
ACTIVIDAD ENZIMATICA
ELECCION DE FRACCIONES
ACTIVIDAD ENZIMATICA
CROMATOGRAFIA DE LIQUIDOC DE ALTA PRECION (HPLC)
ACTIVIDAD ENZIMATICA
ELECTROFORECIC

METODO EXPERIMENTaL
AISLfiMIENTO.
La actinidina fue purificada como un derivado inactivo del tetrationato de la enzima, para prevenir la autodigestión. ( 3 )
2 kg de fruto de Kiwi fueron pelados y licuados durante 30 minutos, con un litro de regulador de fosfatos 10.1 M ) pH = 6.0 conteniendo tetrationato de sodio 1 0 mM y EOTA 1 mM. La mezcla resultante se centrifug6 30 min a 5 000 rpni y 4 'C. El precipitado fue desechado.
PRECIPITaCION.
Del sobrenadante se precipitb la enzima, con sulfato de amonio al 70% de saturaclbn ( 4 8 hrs). El precipitado fué separado por centrifuqaci6n durante 30 min a 5 Oü0 rpm y 4 'C.
*. El precipitado fue resuspendido en el regulador de fosfatos
y dializado frente a agua durante 72 hrs a 4 )C." E.1 solido resultante se redisolvib en la minima cantidad del regulador de fosfatos. ~ . a muest.ra se centrifugó 20 Bin. a 4 000.orpm y 4 "C.
E l nuevo sobrenadante con el extracto crudo de la enzima se almaceri6 a - 20 ' C .
?".' . . ..~. __._I

. , , , ~ . ~ .... .,.. .._~.".I__._._..,".
I
111 1 a I I
FILTRACION EN QEL.
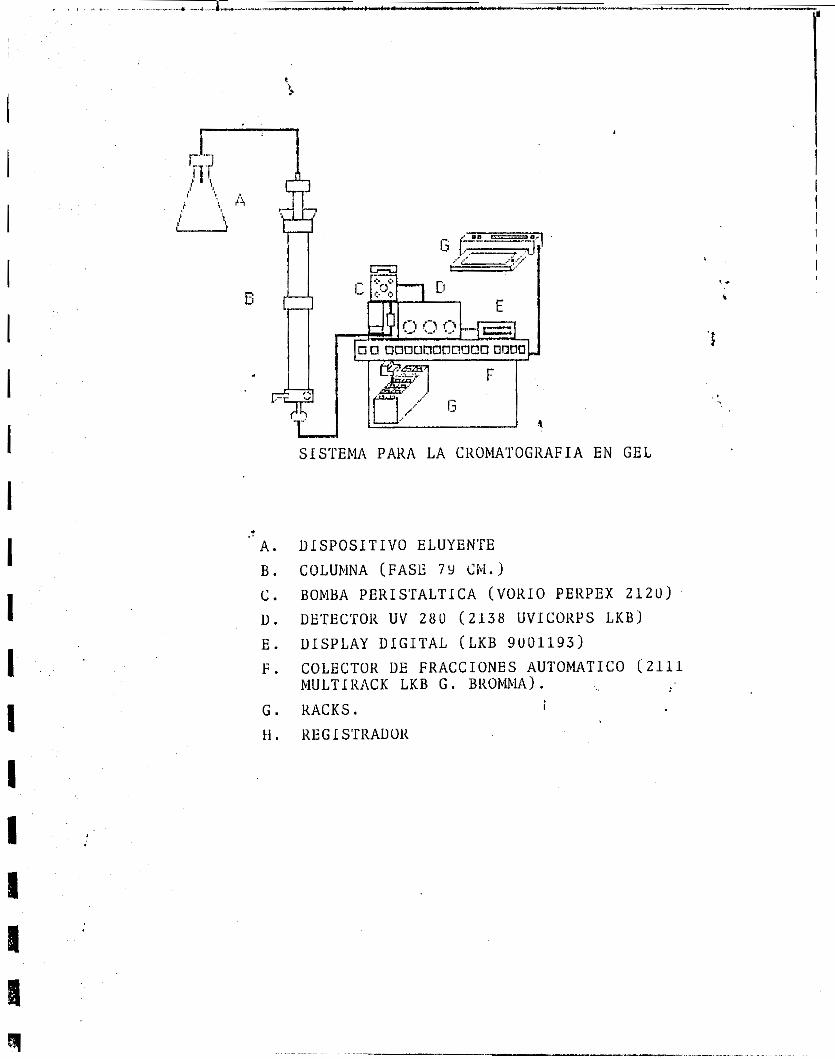
La filtracidn en gel se realizb en una columna d e 79 cm de altura por 2.5 cm de diámetro, empacada con ultrogel FICA 54 y acoplada a un colector automático de colecciones LKB (2111 rlultirac y un detector modelo 2138 Unicord '3). El eluyente utilizado fué regulador de fosfatos 0.1 M pH = 6.0 ( Figura A.)
. .
INTERCAMBIO IONICO.
El intercambio iónico se realizó en una columna de 17 cm de altura por 1.7 cm de diámetro con trisacryl M DEFIE, acoplada a un colector automático d e fracciones LKB. L a eluci6n se hizo utilizando un gradiente lineal de cloruro de potasio de 0.1 a 1.2 M en el regulador de fosfatos (350 ml totales).
CROMATOGRAFIA LIQUIDA DE ALTA PRESION (HPLC)
I 81 81 U
Se utilizó una columna de intercambio cationic0 Bio-gel DEFIE 5-PW de 7.5 cm de altura y 0.75 cm de diámetro acoplada a un cromatógrafo Varian 5020. La elución de la enzima se realizó con un gradiente salino de cloruro de sodio de O a 0.5 Pi en regulador
-. de acetatos 0.05 M pH = 4.0. Se hicieron pruebas de elución con diferentes barridas de gradientes salinos. ( Figura B . )
I
I I I I I I I I I I I I I I 4 81
a
- n .-,
:- . ' I
. I
SISTEMA PARA L A CKOMATOGKAFIA EN GEL
.t A. B. c . u . E. F.
G . H .
D I S P O S I T I V O ELUYEN'I'E
COLUMNA (FASE 7 Y Chi. BOMBA PEKISTALT ICA (VORIO PEKPEX 2 1 2 0 )
DE'rECTOR UV 280 (2138 UVICOKPS LKBJ UISPLAY D I G I T A L (LKB 9001193 ) COLECTOK DE FRACCIONES AUTOMATIC0 (2111 MULTíKACK LKB G . BKOMMA) .
i RACKS.
KEGI S'TRADOK
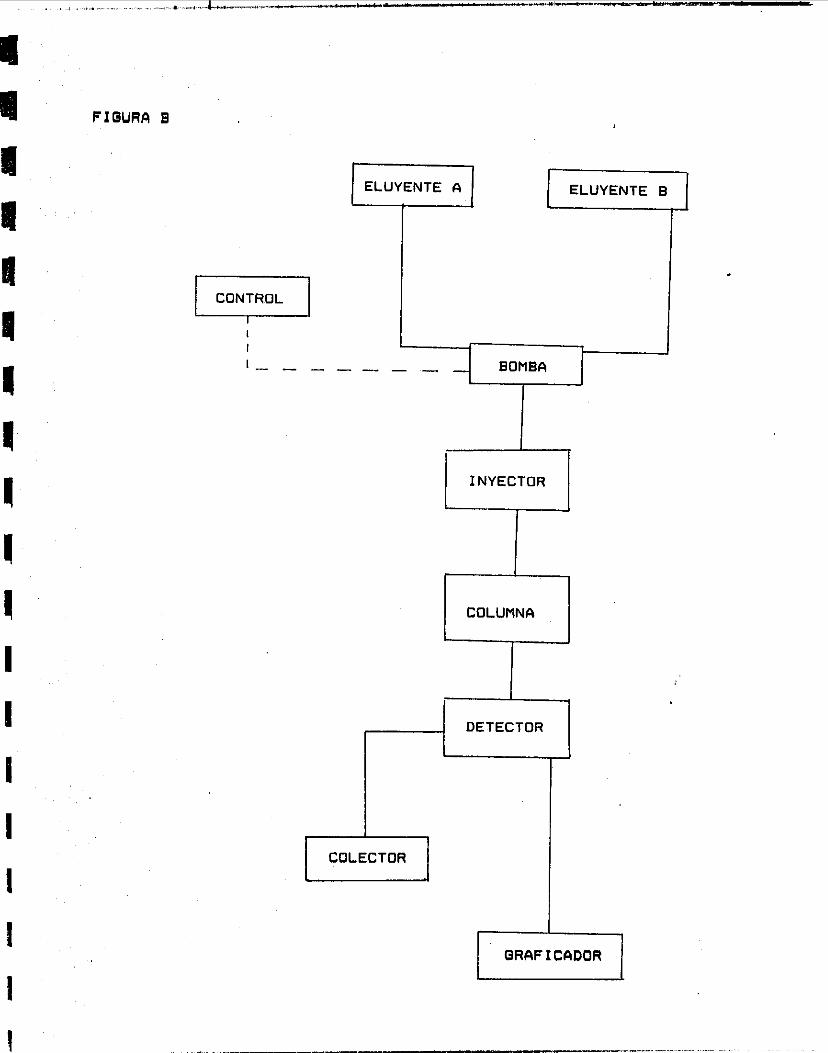
FIOURR B
a B
1 I rl 1 1 1 I
ELUYENTE A ELUYENTE B
E3 COLECTOR
r
COLUMNA
I QRRFICFIDOR 1

ñCT I V IDñD
Se h i c i e r o n pruebas para ob tener l a s cond ic iones 6p t imas para l a medic i6n de l a a c t i v i d a d enz ima t i ca . Se prob6 como s u s t r a t o caseína a l 2 % en peso a pH = 3.8 y 5.81, a s í como hemoglobina a l 2% en peso a pH = 6.13.
Y a que e l s i t i o a c t i v o de l a enzima fué bloqueado du ran te l a e x t r a c c i ó n , con e l t e t r a t i o n a t o de sod io , pa ra p r e v e n i r l a a u t o d i g e s t i b n , para poder medi r l a a c t i v i d a d fue n e c e s a r i o u t i l i z a r un a c t i v a d o r de l a enzima, es d e c i r , desbloquear e l s i t i o a c t i v o .
Se e l i g i e r o n EDTA y C i s t e í n a como a c t i v a d o r y se probaron d i f e r e n t e s concent rac iones y t iempos de i n c u b a c i d n con l a enzima a 37 y 40 oc .
Se h i c i e r o n l a s s i g u i e n t e s pruebas:
a ) EDTA 0.01 M + C i s t e í n a 0.01 M con 1 m i n de i ncubac i6n a 37 OC u t i l i z a n d o coma s u s t r a t o caseina, pH = 3.B. No se observó a c t i v i d a d .
b ) EDTA 0.01 M + C i s t e í n a 0.01 M can 1 m i n de i ncubac i6n a;'37 OC u t i l i z a n d o como s u s t r a t o caseina, pH = 5.81 Se observb l i g e r a a c t i v i d a d .
c ) EDTA 0.01 M + C i s t e í n a 0.01 M con 1 m i n de i ncubac ion a 37 OC u t i l i z a n d o como s u s t r a t o hemoglobina, pH = 6.13 Se observo l i g e r a a c t i v i d a d .

, - , ... ”. .__I ._ . . _.
De las pruebas anteriores de la que mejor resultados se obtuvieron fue la (b), por lo que se eligió para variar las concentraciones de EDTA y Cisteina, así como el tiempo de incubacibn. F.
d) Cisteina 0.01 M y EDTA 0.01 M, tiempo de incubación 1 rnin a 37 oc. Ligera actividad.
.i
e) Cisteina 0.01 M, 1 min de incubación a 37 O C .
Se observó ligera actividad.
f) Cisteina 0.01 M, 1 min de incubación a 40 OC. Se observó ligera actividad.
*. g ) Cisteína 0.001 M, con tiempos de incubacibn de
‘C. No se observb actividad. 1 y 3 min a 40
4 Por lo tanto, de las pruebas anteriores se eligió:
- Sustrato: Caseina al 2 Y. en peso pH = 5.81 - Activad0ra:Cisteina 0.01 M con un tiempo de incubación de 1 min. - Temperatura de reacción: 40 o C.
Por lo tanto, l a actividad proteolítica se midib frente a caseína al 2 7. pH = 5.81, preparada con regulador de fosfatos 0.1 M (pH final 5.81). El activador utilizado para .la enzima fue Cisteina 0.01 M preparada en regulador de fosfatos v.l 11 pH = 6.0
I . . . -

PRUEBA DE ACTIVIDnD.
Al lcuotas de 6 ü O p l de l a enzima + 1 5 0 ~ 1 de act ivador se incubaron durante 1 min a 40 OC. Se aRadieron 1000 pl de l sust ra to . La mezcla resu l tan te , se incubo durante 10 m i n a 40 o "C. La r eacc i6n . se detuvo con 2000p l de bcido t r i c l o r o a c é t i c o al 5 7.. E l prec ip i tado r esu l t an te se separb por centr i fugac i6n a 500 rpm durante 10 min, seguido de una f i l t r a c i ó n a t raves de- papel Whatman de l no. 40. A l f i l t r a d o se l e ley6 absorbancia a 280 nm.
- L a unidad de ac t i v i dad por my a 4CB C se def ine cor~c:
RESULTADOS Y DISCUSION.
PURIFICACION.
FILTRACION EN GEL
En l a figura 1 se presenta e l cromatoyrama de e luc ion par-a l a t i l t r a c i ó n en y e i d e l extracto crudo de Ac t in id ina .

Se observan c u a t r o p i c o s de abso rc ión a 260 n m , de i o 5 cua les solamente e l p i c o no. 2 presenta a c t i v i d a d p r o t e o l i t i c a f r e n t e a caseina. E l p i c o no. 2 f u é concentrado en una s o l a f r a c c i ó n y se rec romatog ra f i o ob ten iendo un s o l o p i c o . La muestra F2,se concent ró pa ra pruebas de i n t e r c a m b i o i 6 n i c o .
INTERCWBIO IONICO.
En l a f i g u r a no. 2 se presenYa e l cromatograma de l a f r a c c i ó n F2 para e l i n t e r c a m b i o i o n i c 0 en columna ( t r i s a c r y l M DEGE), y l a a c t i v i d a d f r e n t e a case ina pa ra cada f r a c c i ó n co lec tada. Se observa que e l máximo de a c t i v i d a d no corresponde a l m a x i m 0 de absorbancia a 280 n m , l a s f r a c c i p n e s que presentaron a c t i v i d a d ( 15 a 19) se concent ra ron en una s o l a f r a c c i ó n : se d i a l i z a r o n f r e n t e a r e g u l a d o r de a c e t a t o (0.05 M) pH = 4.0, du ran te 4 8 h r s . La f r a c c i ó n r e s u l t a n t e 8 K 8 , fue u t i l i z a d a para l a p u r i f i c a c i ó n en HPLC.
a
U
CROMATOGRAFIA L I Q U I D A DE ALTA PRESION (HPLC) la
En l a f i g u r a 3 se presenta e l cromatograma de e l u c i ó n pa ra el i n te rcamb io i ó n i c o en HPLC, se observan 5 p i c o s de absot-ción a 280 nm, de l o s cua les e l p i c o no. 3 (K3) y e l p i c o no. 5 (K5) presentaron a c t i v i d a d p r o t e o l l t i c a f r e n t e a caseína: f r a c c i 6 n K3, 216 U/mg y f r a c c i ó n K5, 120 Uimg.
ELECTROFORESIS
Las pruebas de e l e c t r o f o r e s i s muestran una s o l a banda para l a enzima K3 y una s o l a banda para l a enzima K5.

F 2
l i 1 0 20 30 4 0 50 60 70 80 ~ r a c c i ó n .
¡gura 1.

1 1 I I 1 I I I I I I I 1 1 B
!I
0 8 K C 1 L 1 . 2 M I O rl O
O
I
. Actividad t U / m g 1
O O O O O O
io oio cu
\
I I I I
O cv
io r
C *O o o tu LL
.-
L
O r
io
O
U tu U > o . a
.-
.- +I
+ O o C .O
O
.-
.-
.- n E a o
. a 2 C
L
' + I - I

, "
I I I I I I I II a II B a B 8
71
I I I I I I
t
.

e 1
Ba i
CONCLUSION
Se o b t u v i e r o n puras dos formas molecu la res de l a a c t i n i d i n a , d e l f r u t o d e l K i w i ( a c t i n i d i a c h i n e n s i s ) , l a s cua les no habían s i d o repo r tadas an te r io rmen te , es tab lec iendo l a r u t a de p u r i f i c a c i ó n y las cond ic iones 6p t imas para de te rm ina r l a a c t i v i d a d p r o t e o l í t i c a de ambas enzimas.
PERSPECTIVñS.
Una vez que se han p u r i f i c a d o l a s enzimas, se i n i c i a r a n 105 e s t u d i o s e s t r u c t u r a l e s po r d i c r o i s m o c i r c u l a r . E s t e t i p o de e s t u d i o s se r e a l i z a r á n con e l o b j e t i v o de una c l a s i f i c a c i 6 n e s t r u c t u r a l de l a enzima, s i n t e n e r necesidad de r e c u r r i r a e s t u d i o s de d i f r a c c i ó n de rayos X .

REFERENCIAS.
- ( 1 ) GLAZAR, A. N. & SMITH, E. L. (1971) IN THE ENZYMES, P.D BOYER, Ed. 3era. ed. vol. 3 Academic Press.. new York, pp. 501- 546.
- ( 2 ) BAKER, E.N. & DRENTH, J . (1987) IN BIOLOGICAL MACROMOLECULES ADN ASSEBLIES; Jurmak, F. A. & McPherson, A., Editors. Vol 3. John Wiley & sons. New York pp. 313-368.
- ( 3 ) BROCKELHURCT, K. (1982) METHODS ENZYMOL. 87C 427-469
- ( 4 ) BAINES, B.C.& BROCKLEHURCT, K., (1982) BIOCHEM J. 205 205-21 1
- ( 5 ) CARNE, A. & MOORE, C. H (1978) BIOCHEM Ji.173, 73-83
- (6) BROCKELHURCT, K. BAINEC, B.C., & KIECRTAN, M. P., (1981) . TOP ENZYME FERMENT BIOTECHNOL 5 262-335.
- ( 7 ) HUSAIN, C. S & LOWE, G. (1970), BIOCHEM J . 116, 689-692
- (8) DRENTH, J., HOL. W. G., VICSER, J . W. E. & SLUYTERMAN, L.A.A.E., (1968) J . MOL BIOL. 34 369-371.





























