Embed Size (px)
Citation preview

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 1 af 31
1. SOMATOSENSORISK FUNKTION = huden som sanseorgan
Disposition: Stikord
1. Somatosensoriske sansemodaliteter Taktil, temperatur, smerte, proprioception 2. Taktile receptorer Pacini, Ruffini, Meissner, Merkel 3. Signalet og signalmodulation Alle stimuleres samtidigt, lateral inhibition 4. Ascenderende taktile baner Kroppen og ansigtet, 3 neuroner, somatopi 5. Repræsentation i cortex Homunculus gentages - dog mere integreret, plasticitet
1. Somatosensoriske sansemodaliteter: Ordnet efter adækvat stimulus: a) Tryk, berøring og vibrationer (Mekanoreceptorer). ”Den taktile sans”. Dette emne. b) Temperatur (Thermorecptorer) – se emne 2 c) Smerte (Nociceptorer) – se emne 2 d) Stilling (Proprioceptorer) – se emne 7 Skal vha. nerveimpulser give rapport til hjernen om hudens omgivelser mht: Hvad der stimuleres med samt hvor, hvor meget og hvornår der stimuleres Receptive felt = det område i omverden som ændrer et givent neurons membranpotential 2. Taktile receptorer: Mekanosensitive (Na+) kanaler i nerve terminal (axon!) Åbner ved deformering (tryk, berøring, vibrationer, stræk) Hvis nok kanaler depolariserer på en gang = aktionspotential Tæthed af receptorer varierer (tættest ved fingerspidser og læber, jf. topunktsdiskrimination) Forskellige mekanoreceptorer: Navn Lokalisering Receptivt felt Adækvat stimulus Adaptation Pacini legemer Dybt Stort, samlet 200-300 Hz Hurtig Ruffini ender Dybt Stort, samlet < 50 Hz Langsom Meissners legemer Superficielt Små, spredte 50 Hz Hurtig Merkels diske Superficielt Små, spredte < 50 HZ Langsom
huskefif: ”De italienske ligger dybere/sydligere end de tyske!” ”Meissners adaptation er schnell!” ”Pacinis kapsel absorberer tryk og er derfor også hurtig til at adaptere!” Skal kunne reagere på fare → impuls føres i hurtige Aβ axoner (40-80 m/sek). 3. Signalet og signalmodulation: Receptor er mest følsom lige ved receptorlegemet Alle typer receptorer stimuleres normalt på en gang: • Dybe vs.overfladiske Tidligt et nuanceret og informativt signal • Hurtigt vs. langsomt adapterende • Forskellige receptive felter (overlap). • Lateral inhibition se nedenfor

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 2 af 31
Ved hver relæstation (nucleus cuneatus, thalamus, cortex) moduleres signalet endnu mere: • Receptive felt bliver større og større / information spredes mere og mere (et neuron i cortex
snakker med mange forskellige receptor-legemer. Lateral inhibition: En receptor har inhibitoriske interneuroner til naboneuroner Når receptoren påvirkes sender den:
a) et stærkt excitatorisk signal i sin egen ascenderende synapse b) et mindre inhibitorisk signal til nabocellernes ascenderende synapser
effekt = større kontrast mellem der hvor huden stimuleres og ikke stimuleres. topunktsdiskriminationen øges. kan benyttes til (mere præcist) at føle en genstand der bevæger sig over huden. 4. Ascenderende forløb: 3 neuroner til cortex. Taktil information fra kroppen ascenderer i bagstrengsbanen, dvs. • 1. neurons cellelegeme er placeret i spinalgangliet • Ascenderer uden synapse ipsilateralt i fasciculus gracilis ell. cuneatus (i funiculus posterior) • Første synapse i nucleus gracilis og cuneatus (i medulla oblongata) • 2. neuron krydser her midtlinjen (decussatio lemniscorum) og ascenderer i lemniscus medialis • Når nucleus ventralis posterolateralis (VPL) i thalamus og synapser her • 3. neuron ascenderer til somatosensorisk cortex Taktil information fra ansigtet ascenderer sideløbende med lemniscus medialis, dvs. • 1. neurons cellelegeme ligger i nucleus trigeminale og synapser i nucleus trigeminale pontinus. • 2. neuron krydser her midtlinjen og ascenderer i lemniscus trigeminale • Når nucleus ventralis posteromedialis (VPM) i thalamus og synapser her • 3. neuron ascenderer til somatosensorisk cortex Fibre føres altid somatotopisk (nærliggende områder føres sammen) Dvs. somatopi både i baner og relæstationer (ved hver synapse) Arealet for hvert område på kroppen er proportionalt med tætheden af receptorer (jf. homunculus) 5. Repræsentation i cortex: Primære somatosensoriske cortex:, S1: Brodmans område 3a, 3b, 1 og 2 i gyrus postcentralis Homunculus: ”står på corpus callosum” (dvs. hovedet er mest lateralt) Homunculus gentages i hvert område (samme stimulus afbilledes i hvert område) Mere sofistikerert jo længere ”bagud” man kommer. • 3a og 3b kan vurdere størrelse af objekter. • 1 kan vurdere hvor overfladen af objekter. • 2 kan vurdere retningen af et objekt der flytter sig over huden. • Signal sendes videre til S2 (område 5 og 7) som kan vurdere input bilateralt. ”samler signal” Søjlearrangement i cortex: en søjle modtager f.eks. kun hurtigtadapterende signaler. Receptive felter overlapper.

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 3 af 31
Input sendes til lag IV (lamina granularis interna) Output kommer fra lag V (lamina pyramidalis interna) Plasticitet i CNS: Område i cortex kan blive større og mindre. Studier af aber med: • Amputeret 3. finger (2. og 4. fingers områder i cortex tager over) • Overstimulation af 3+4. finger (3+4. fingers områder bliver større) Kroniske fantomsmerter: • Amputeret legemesdels innervationsområde i cortex overtages af smerteområde.

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 4 af 31
2. SMERTE Disposition: Stikord
1. Definitioner af smerte nociceptiv vs. neuropatisk 2. Nociceptorer mekano, thermo, elektro, kemo, pH + polymodale 3. Ascenderende smertebaner adskilt fra taktile baner, krop vs. ansigt 4. First og second pain Aδ fibre til cortex vs. C fibre til limbiske system 5. Sensibilisering og regulering 1 x perifær-, 2 x central smerteregulering. 6. Refereret og projiceret smerte angina pectoris vs. fantomsmerter 7. Eksempel på en smerterecptor capsaicin (chili)
1. Definitioner af smerte Smerte: den ubehagelige sensoriske og emotionelle oplevelse, der opstår når væv skades eller lider overlast (…) eller oplevelser der beskrives i tilsvarende vendinger! (IASP) Nociceptiv smerte = Smertereceptorer aktiveres Neuropatisk smerte = Det der føles i hjernen 2. Nociceptorer: Adekvat stimulus: ”fare for vævsbeskadigelse” Forskellige undergrupper: a) mekaniske påvirkninger (mindre følsomme end alm. trykreceptorer) b) ekstreme temperaturer (over 45 °C, under 20 °C) c) elektrisk stimulering d) kemiske substanser (ex. histamin fra mastceller) e) pH (H+ sensitive, feks. karskade → ÷ iltforsyning → laktat) a + b + c er simple frie nerveender uden en omgivende kapsel d + e aktiveres indirekte vha. specielle membranproteiner! Nogle receptorer virker på mere end blot en type stimuli = polymodale Virker parallelt med andre modaliteter, f.eks b) som virker parallelt med normale thermoreceptorer: Thermoceptorer føler varmt Temperaturfølsomme nociceptorer føler brændende (smerte er en separat sans!)
3. Ascenderende smertebaner: Følger andre baner til cortex end taktile stimuli → cortex kan skarpt adskille forskellige stimuli. Stor integration af stimuli ved hver synapse. Ascenderer kontralteralt
ene afhænger ikke nødvendigvis af den anden

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 5 af 31
Kroppen: 1. neurons cellelegeme ligger i spinalgangliet Danner synapse i baghornets substantia gelatinosa (ascenderer evt. et enkelt segment først) 2. neuron krydser langs commisura alba (foran canalis centralis) Ascenderer i den tractus spinothalmicus imellem funiculus anterior og lateralis Danner synapse i VPL thalamus
Ansigtet: 1. neurons cellelegeme ligger i ganglion trigeminale Løber til medulla oblongata niveau ender i nucleus spinalis n. trigeminus 2. neuron krydser midtlinjen og løber til VPM thalamus (i lemniscus trigeminale) Smertebaner terminer forskellige steder i thalamus (dvs. separat fra taktile baner) Krydsning sker på andet niveau end taktil → dissocieret sansetab ved halvsidig tværsnitslæsion Mange interneuroner undervejs indgår i smertekontrol (se nedenfor) og refleksbuer 4. First og second pain: First pain: • Den umiddelbare og præcise smerte når man slår sig • Ledes af Aδ fibre (dvs. relativt hurtigere – omk. 20 m/sek) • Skærende og præcis smerte • Baner løber fra thalamus til somatosensoriske cortex (stadig adskilt fra taktile områder) Second pain: • Diffus • C fibre (umyeliniserede, dvs. langsomme – omk. 2 m/sek) • Trykkende og upræcis smerte • Påvirker blandt andet det autonome nervesystem (smerten medinddrager hele kroppen) • Baner løber fra thalamus til det limbiske system og mange kommer ikke til bevidstheden 5. Sensibilisering og regulering: Perifær sensibilisering: • Nociceptorer sender ”afstikkere” som udskiller substans P vævets mastceller → histamin →
a) vasodilation/inflammation b) sensibilsering af vævets andre nociceptorer =hyperalgesi
• F.eks hyperalgesi når en solbrændt hud er meget følsom. • Kaldes også for axon-refleksen (involverer kun et neuron!) Central smerteregulering (1): • Inhibitorisk interneuron fra sensorisk til nociceptive neuron. • Kun nociceptor = maximal fyringsfrekvens • Også sensoriske neuron = maximal inhibering → smertesignal hæmmes • Derfor man ryster hånden hvis man slår sig med en hammer

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 6 af 31
Allodyni = Når taktil berøring kommer til at opfattes som smerte. En sygelig og kronisk tilstand som måske skyldes at mekanismen ovenfor skades. Central smerteregulering (2): • I stresset situation kan smerte-signal undertrykkes • Periakvæduktelle Grå Substans (PAG) aktiveres i mesencephalon (aktiveres af limbiske system) • Synapse i nucleus raphes magnus (NRM). • Serotonerg synapse med interneuron i baghornet • Interneuron udskiller enkephaliner som påvirker opiate receptorer på det præsynaptiske
smerteneuron (C-fiber) i baghornet.
• Eksempelvis soldater og fødende der påvirker deres smertetærskel (hypoalgesi) • Enkephaliner altså dannet af kroppen selv • Opiater er farmaka hvis virkning er tilsvarende enkephaliners. NB: opium har andre virkninger
andetsteds i kroppen. 6. Refereret og projiceret smerte: Refereret smerte: Visceral smerte er sjælden, somatisk smerte er hyppigere. Ofte ”deler” viscerale og somatiske nociceptorer 2. neuron → smerte fejlfortolkes af hjernen Ses eksempelvis ved angina pectoris (n. phrenicus og nn. supraclaviculares deler C4) Projiceret smerte: Uanset hvor en nerve blev stimuleret vil hjernen ”projicere” impulsen ud til nervens udspring
Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 7 af 31
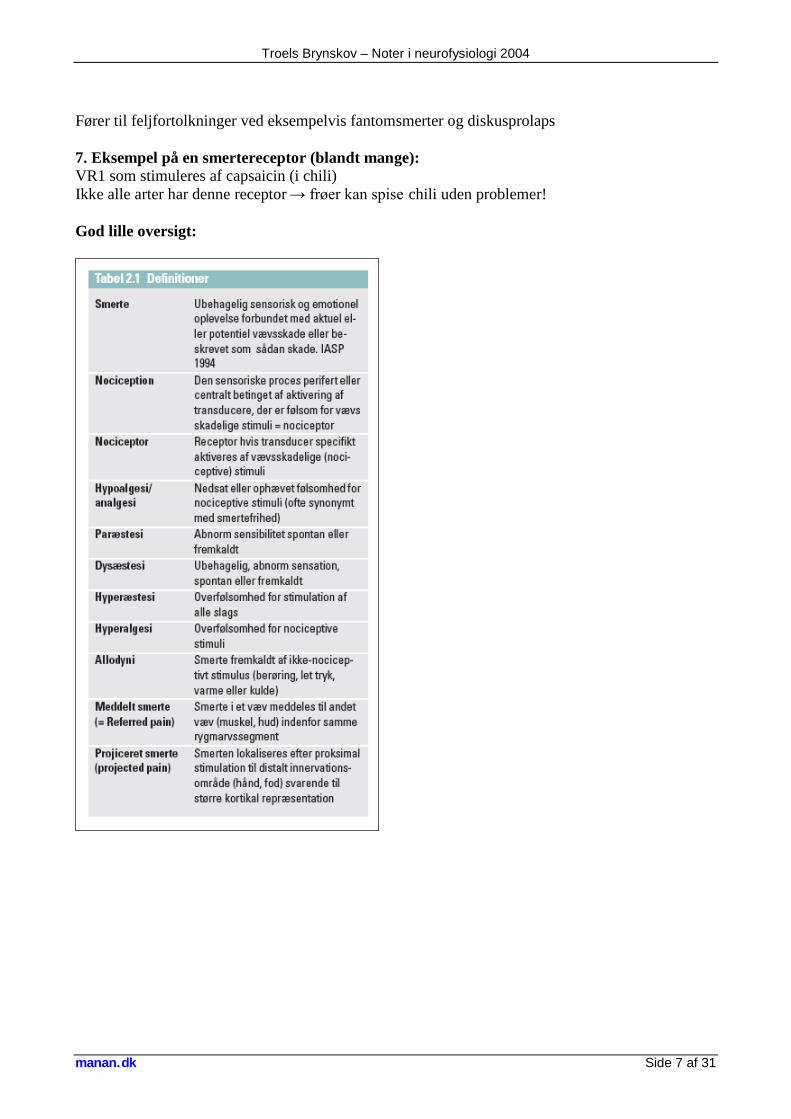
Fører til feljfortolkninger ved eksempelvis fantomsmerter og diskusprolaps 7. Eksempel på en smertereceptor (blandt mange): VR1 som stimuleres af capsaicin (i chili) Ikke alle arter har denne receptor → frøer kan spise chili uden problemer! God lille oversigt:

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 8 af 31
3. ØJET Fra det øjeblik lys rammer retina og til output sendes fra gangliecellerne
Disposition: Stikord
1. Øjets funktioner syn, balance, døgnrytme 2. Oversigt over retina 3 cellelag, lys passerer andre lag til fotoreceptorer 3. Fotoreceptorer tappe vs. stave, opsin vs. retinal, transduktion 4. Ganglieceller M: bevægelse, P: blå/rød, form og detalje, nonM-NonP: gul/blå 5. Synaptiske mekanismer imellem fotoreceptorer og ganglieceller Bipolære celler: ON respons (ionotrop) vs. OFF respons (metabotrop) 6. Kant-detektering ON-center vs. OFF-surround.
1. Øjets funktioner: Synsperception: Dagssyn/farvesyn (fotopisk syn) Nattesyn (skotopisk syn) Automatiske funktioner: Orienteringsreflekser (via fasciculus longitudinale medialis) Pupilkontrol Døgnrytme (via nucleus suprachiasmaticus) Balance (sammen med vestibulærapparatet og mekanoreceptorer) 2. Oversigt over retina:
Der findes forskellige undertyper af de enkelte celletyper (ikke pensum) 3. Fotoreceptorer: Sidder inderst mod pigmentcellelaget Stave: Mange diske (→ meget lysfølsomme) 1 slags opsin (rhodopsin – ikke farvefølsomme) Størst tæthed i periferien af fovea centralis, desuden mange i perifære retina. Mættes hurtigt (kan ikke anvendes i dagslys) Tappe: Få diske (→ ikke så lysfølsomme som stave)

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 9 af 31
1 ud af 3 opsiner (Blå, Grøn, Rød – følsomme overfor forskellige bølgelængder = farve). Størst tæthed i midten af fovea centralis. Det molekyle som opsin binder til: ”Retinal”. Dannes fra vitamin D. Tappe og stave er grundlaget for duplex syn = to paralelle systemer. Transduktion ved lys-input: 1 foton → 1 retinal aktiveres → mange phosphodiesterasers (PDE) aktivitet øges (forstærkning af signal) → mange PDE nedbryder mange cGMP molekyler → ”rigtig mange” Na+ kanaler lukker → cellen hyperpolariserer. I mørke dannes flere retinal- og opsinmolekyler → større lysfølsomhed (ca. 30 minutter). 4. Ganglieceller: Eneste som fyrer aktionspotentialer (fotoreceptorer og bipolære celler medierer receptorpotentialer) Magnocellulære: M-type Store. Stort receptivt felt. Ca. 10% af alle ganglieceller. Kan detektere bevægelse Parvocellulære: P-type Små. Små receptive felter Ca. 80% af alle ganglieceller. Kan detektere rød/grøn opponens, form og detaljer. NonM-nonP: Dårligt karakteriserede. Gul/blå opponens (Gul syntetiseres vha. rød- og grønfølsomme fotoreceptorer) 5. Synaptiske mekanismer mellem fotoreceptor og ganglieceller: Beskrives her for M-type (samme princip med farve opponens for P og nonM-nonP):

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 10 af 31
Hver ganglioncelle er enten ON-center eller OFF-center med modsat surround i sit receptive felt. Center bestemmes af de bipolære celler – hvoraf der findes to typer (ON og OFF) OFF-center: Mørke i midte depolariserer fotoreceptor → frisætter glutamat til bipolære celle. OFF-bipolære celles ionotrope kation-kanaler åbner og bliver derved et EPSP. ”Det normale” ON-center: Mørke i midte depolariserer fotoreceptor → frisætter glutamat til bipolære celle.
ON-bipolære celles metabotrope G-protein receptorer lukker Na+ kanaler → medierer IPSP. ”Usædvanligt” (glutamat som inhibitorisk transmitter)
Surround bestemmes af de horisontale cellers laterale inhibition Surround: Horizontale celler påvirkes og inhiberer den bipolære celle (nt = GABA). Hvordan de nogle gange laver OFF og nogle gange ON er ikke endeligt afklaret NB: En enkelt fotoreceptor kan altså indgå som OFF-center og ON-center alt afhængig af de celler som den danner synapse med. Dette kaldes parallel processing. Sammenfatning: Fotoreceptor omkoder mørke hhv. lys og frisætter hhv. blokerer glutamat frisættelse. De bipolære celler som fotoreceptoren synapser med har enten et OFF eller et ON respons. De bipolære cellers synapser med gangliecellerne sørger for at center bliver forskelligt fra surround. Kant-detektering: Center vs. surround antagonist → stor følsomhed overfor kanter:
Forskellen i lys/mørke refleksion bestemmes på denne måde relativt (ikke antal fotoner pr. areal)

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 11 af 31
4. CENTRALE SYNSMEKANISMER Dispostion: Stikord
1. Forbindelser fra retina cortex, FLM, tectum, hypothalamus, 2. Celletyper i synsabarken orienterings-, retnings- og komplekse celler 3. Lag i synsbarken IV (retning), blob (form) og interblob (farve) 4. Forbindelser fra V1 og V2 videre i cortex. V3-4-5, dorsal/ventral strøm, afstandsbedømmelse.
1. Forbindelser fra retina (retinofugal projektion):
A. Corpus geniculatum laterale (videre til synscortex). B. Colliculus superior (øjenenes bevægelser integreres med andre stimuli). C. Tectum (lysrefleksen – pupilsammentrækning) D. Hypothalamus (døgnrytmer) E. Amygdala (limbisk respons)
Corpus geniculatum laterale: Banerne forløber fra retina i det ”klassiske forløb” hvor den nasale retina krydser i chiasma opticum.
I corpus genicucaltum laterale ses 6 adskilte lag. • 2 inderste: Magnocellulære system (et lag fra højre og venstre side) • 4 yderste: Parvocellulære system (to lag fra højre og to lag fra venstre side) • Indimellem de andre lag: Koniocellulære system • Man kan evt. huske at de yderste lag er fra den kontralaterale side! Der ses retinotopi, bemærk at: • Billedet vendes på hovedet og spejlvendes • Foveas område er stærkt forstørret • Receptive felter i corpus geniculatum laterale svarer til retina, ca. 1º (som månen!). • Øjets store opløsning fås ved at de mange receptive felter overlapper hinanden. Forbindelse til synscortex (V1): • Fibre forløber via radiatio optica • Mange projektioner fra fovea centralis → går ofte fri ved delvis ensidig parese i radiatio optica

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 12 af 31
• Der sendes corticofugal feedback fra synsbarken til corpus geniculatum laterale.Funktion heraf er dog uvist – grunden til de lysglimt man oplever i mørke?
Colliculus superior (også tectum?): • Integrerer auditive, taktile og visuelle stimuli • Alle tre modaliteter er ”topisk” ordnet (dvs. retinotopt for øjet) • Colliculus superior er medvirkende til orienteringsrefleksen via FLM) Tectum: • Fibre fra retina løber til visuelle reflekscentre i tectum bilateralt • Det afferente led af puilrefleksen (hvor begge pupiller kontraherer sig ved lys stimuli) Hypothalamus: • Fibrene til nucleus suprachiasmaticus er med til at regulere det indre ur • Receptorer er specialiserede ganglieceller (ikke fotoreceptorer!) • Behandles videre under ”hjernens rytmer” Amygdala: • Forsøg med konduktivt blinde der kan registrere følelsesprovokerende ansigter! 2. Celletyper i synsabarken: Orienteringsselektive celler (simple celler): • 1 medial aktiverende zone (i dette eksempel med ON-center ganglieceller) • 2 laterale inhiberende zoner • Maksimalt stimulus når lysstråle falder i den mediale og ikke i de laterale zoner
Retningsselektive celler: • Binokulære • Kan detektere retningen af stimlus • F.eks. mere følsomme hvis stimulus kommer fra venstre mod højre end omvendt • Har store receptive felter på op til 10º

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 13 af 31
Komplekse celler: Meget mere orienteringsfølsomme end simple celler. 3. Lag i synsbarken: Primære synscortex ligger i den mediale occipitallap omkring sulcus calcarinus Kaldes enten Brodmans area 17 eller V1. Lag IV: Modtager al synsinput (lag IV meget stort i V1) Mange orienterings- og retningsselektive celler (jf. ovenfor) → analyse af bevægelse Ordnet i okulær dominans striber Højre- og venstresidigt input holdes adskilt i et zebra (eller ”pinwheel”) mønster Magnocellulære system ender her. Lag II og III: Blander input fra begge øjne Parvocellulære system ender her. Blobs: Mange komplekse celler (jf. ovenfor) → analyse af form Interblobs: Analyse af farve 4. Forbindelser fra V1 og V2 videre i cortex: Groft sagt integreres stimuli videre i synscortex til: • V3: Dynamisk form (formen på objekter i bevægelse). • V4: Farve-opfattelse. • V5: Bevægelse. Og dernæst videre i: • En ventral informationsstrøm til temporallappen med detaljer og farver. • En dorsal informationsstrøm til parietallappen med grove konturer og hurtige bevægelser. The binding problem: information spredes og spredes, men hvordan samles den til bevidstheden. 3-D og afstandsbedømmelse:

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 14 af 31
Monookulære mekanismer: • Perspektiv (linjer konvergerer mod horisonten) • Objektkendskab (en stor ting er tættere på end en lille ting) • Interposition (en ting der dækker over en anden må være tættere på) • Parallakse (ting der er fjerne bevæger sig langsommere over horisonten) • Lys/skygge (”lys kommer som regel oppefra”) Binokulære mekanismer (stereoskopisk syn): • Disparate celler kan vurdere hvordan objekter er placeret ift. genstand i fokus.

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 15 af 31
5. HØRESANS Disposition:
1. Lydens vej fra ydre til indre øre 2. Hårcellernes transduktionsmekanisme og vejen til cortex 3. Kodning af frekvens og amplitude 4. Retningsbestemmelse 5. Centrale hørebaner 6. Klinik
Lyd = meget små fortykninger/fortætninger i luft (lydbølger) Fysiologiske problem: at overføre svingninger i luft til nerveimpuls 1. Lydens vej fra ydre til indre øre: Ydre øre: Vibrationer (lyden) passerer følgende strukturer: • Øremuslingen (vibrationer i luft) • Meatus acusticus externus (vibrationer i luft) • Trommehinden (vibrationer i membran) Funktion: Indsnævring af tværsnitsareal → (? x) forstærkning af signal Mellemøret: Vibrationer passerer følgende strukturer: • Trommehinden (vibrationer i membran) • Malleus, incus og stapes (vibrationer af knoglekæde, stapes ”vipper” på det ovale vindue) Funktion: a) Overførsel af svingninger i luft til vibrationer i væske
b) Indsnævring af tværsnitsareal (x 10) og vægtstang (x 2) → x 20 forstærkning af signal
Desuden: Eustachiske rør (trykudligning) Indre øre: Vibrationer passerer følgende strukturer: • Ovale vindue (vibrationsoverførsel fra stapes til perilymfe) • Perilymfen i scala vestibuli (vibrationer i væske) • Perilymfen i scala tympani (vibrationer i væske) • Runde vindue (svingninger i membran) Imellem scala vestibuli og tympani forplantes vibrationer yderligere: • Membrana vestibularis/Reisners membran (svingninger i membran) • Endolymfen i scala media (vibrationer i væske) • Membrana basilaris (svingninger i membran) Der opstår således en travelling wave i membrana basilaris. Desuden: vestibulære system (÷ ovalt vindue for perilymfen at forskyde sig ud i → ÷ vibrationer) 2. Hårcellernes transduktionsmekanisme: Det cortiske organ:
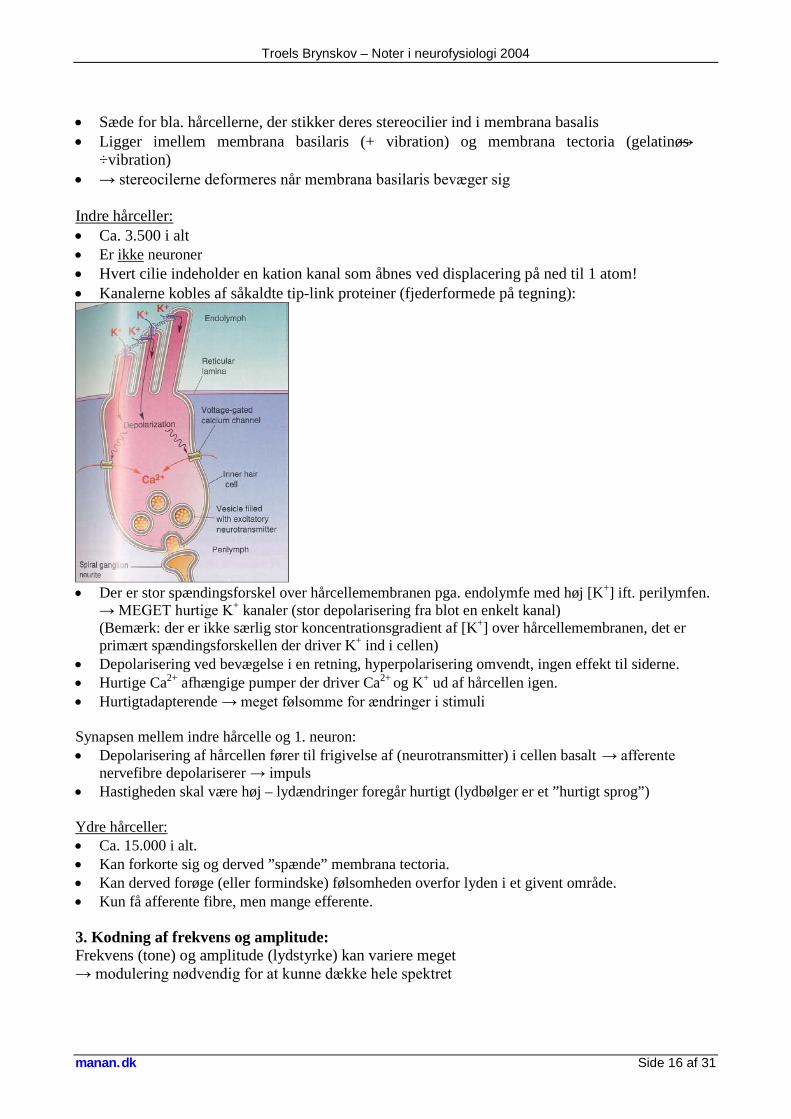
Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 16 af 31
• Sæde for bla. hårcellerne, der stikker deres stereocilier ind i membrana basalis • Ligger imellem membrana basilaris (+ vibration) og membrana tectoria (gelatinøs →
÷vibration) • → stereocilerne deformeres når membrana basilaris bevæger sig Indre hårceller: • Ca. 3.500 i alt • Er ikke neuroner • Hvert cilie indeholder en kation kanal som åbnes ved displacering på ned til 1 atom! • Kanalerne kobles af såkaldte tip-link proteiner (fjederformede på tegning):
• Der er stor spændingsforskel over hårcellemembranen pga. endolymfe med høj [K+] ift. perilymfen.
→ MEGET hurtige K+ kanaler (stor depolarisering fra blot en enkelt kanal) (Bemærk: der er ikke særlig stor koncentrationsgradient af [K+] over hårcellemembranen, det er primært spændingsforskellen der driver K+ ind i cellen)
• Depolarisering ved bevægelse i en retning, hyperpolarisering omvendt, ingen effekt til siderne. • Hurtige Ca2+ afhængige pumper der driver Ca2+ og K+ ud af hårcellen igen. • Hurtigtadapterende → meget følsomme for ændringer i stimuli Synapsen mellem indre hårcelle og 1. neuron: • Depolarisering af hårcellen fører til frigivelse af (neurotransmitter) i cellen basalt → afferente
nervefibre depolariserer → impuls • Hastigheden skal være høj – lydændringer foregår hurtigt (lydbølger er et ”hurtigt sprog”) Ydre hårceller: • Ca. 15.000 i alt. • Kan forkorte sig og derved ”spænde” membrana tectoria. • Kan derved forøge (eller formindske) følsomheden overfor lyden i et givent område. • Kun få afferente fibre, men mange efferente. 3. Kodning af frekvens og amplitude: Frekvens (tone) og amplitude (lydstyrke) kan variere meget → modulering nødvendig for at kunne dække hele spektret

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 17 af 31
Frekvens: • Mennesket kan høre mellem 20 og 20.000 Hz. • Øret er optimeret til mellemområdet (200-4000 Hz). Området for normal tale. • Folk med absolut gehør kan skelne imellem 1000 Hz og 1001 Hz. Membrana basilaris: • Lyden deformerer membranen som når man ryster et lagen: en travelling wave. • Deformeringen af membranen varierer i dens udstrækning pga. dens fysiske egenskaber:
o Stiv og smal ved basis (svinger mest ved høje frekvenser) o Slap og bred ved apex (svinger mest ved lave frekvenser)
• Membranen ”piller” således de enkelte frekvenser ud af lyden. Hårcellerne står for at afkode deformeringen til et signal: • Ca. 10 axoner omkring en hårcelle, men i forskellig afstand fra hårcellen → • Forskellen i mætning af de enkelte axoner afslører inteniteten, altså hvor stor deformeringen
var. • Det område med størst deformering ”vinder” i hjernens tolkning (lateral inhibtion involveret?) Supplerende mekanisme (under 4000 Hz): • Neuroner fyrer ikke ved hver svingning, men ved f.eks. hver fjerde. • Når blot antallet af svingninger som ”hoppes over” kendes kan frekvensen udregnes af hjernen. • Kaldes phase locking (=frekvens kodning?) • Ved frekvenser under 200 Hz kan der laves phase locking ved hver fase. • Sparer neuroner og er sandsynligvis den eneste mekanisme der benyttes under 200 Hz. Amplitude (lydstyrke): • En tidobling i lydamplitude tolkes som en fordobling i hjernen (altså en logaritmisk skala!) • Ved mål for lydstyrke sættes det lige akkurat hørebare (høretærsklen) til 0 (Db) • Ved 120 Db er der fare for skade af hårcellerne • Høj amplitude giver større udsving på membrana basilaris →
a) Større depolarisering af den enkelte hårcele → højere fyringsfrekvens. b) Naboceller stimuleres kraftigere → højere fyringsfrekvens fra celler i området.
4. Retningsbestemmelse: Horisontalt: • Nyankommende lyd (alle frekvenser): Interaural tidsforskel Præcise binaurale neuroner i nucleus olivarius superius modtager impulser fra hver side. Hvert neuron har således sin egen retnings specificitet:
• Konstant lyd (20-2.000 Hz):
faktor 100 forskel!
Højre: Venstre: Signal ankommer først Signal ankommer senere
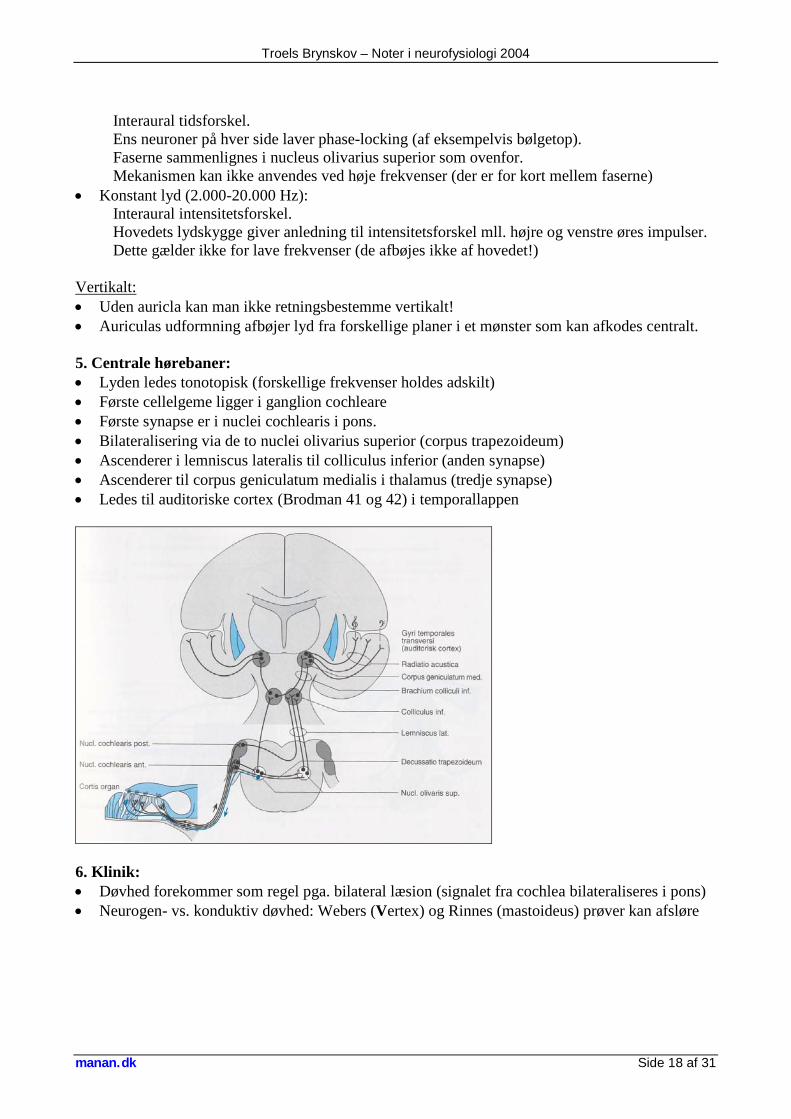
Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 18 af 31
Interaural tidsforskel. Ens neuroner på hver side laver phase-locking (af eksempelvis bølgetop). Faserne sammenlignes i nucleus olivarius superior som ovenfor. Mekanismen kan ikke anvendes ved høje frekvenser (der er for kort mellem faserne) • Konstant lyd (2.000-20.000 Hz): Interaural intensitetsforskel. Hovedets lydskygge giver anledning til intensitetsforskel mll. højre og venstre øres impulser. Dette gælder ikke for lave frekvenser (de afbøjes ikke af hovedet!) Vertikalt: • Uden auricla kan man ikke retningsbestemme vertikalt! • Auriculas udformning afbøjer lyd fra forskellige planer i et mønster som kan afkodes centralt. 5. Centrale hørebaner: • Lyden ledes tonotopisk (forskellige frekvenser holdes adskilt) • Første cellelgeme ligger i ganglion cochleare • Første synapse er i nuclei cochlearis i pons. • Bilateralisering via de to nuclei olivarius superior (corpus trapezoideum) • Ascenderer i lemniscus lateralis til colliculus inferior (anden synapse) • Ascenderer til corpus geniculatum medialis i thalamus (tredje synapse) • Ledes til auditoriske cortex (Brodman 41 og 42) i temporallappen
6. Klinik: • Døvhed forekommer som regel pga. bilateral læsion (signalet fra cochlea bilateraliseres i pons) • Neurogen- vs. konduktiv døvhed: Webers (Vertex) og Rinnes (mastoideus) prøver kan afsløre

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 19 af 31
6. LIGEVÆGTSSANS Disposition:
1. Vestibulærapparatets funktioner 2. Buegangene (canales semicirculares) 3. Sacculus og utriculus 4. Centrale ligevægtsbaner 5. Den vestibulookkulære refleks (et eksempel)
1. Vestibulærapparatets funktioner: Registrerer accelerationer bla. for at vi kan: 1. Påvirke muskeltonus (refleksmæssigt) så vi holder balancen 2. Stabilisere synsindtryk (refleksmæssigt) på fovea centralis Virker sammen med proprioception og synet. (hjernen kan lære at substituere efter læsion) 2. Buegangene (canales semicirculares): • 3 buegange: ossøse labyrint (med perilymfe) med hindede labyrint indeni (med endolymfe) • En ampulla i hver buegang indeholder:
a) En crista ampullaris med hårceller og stereocilier b) En cupula ampullaris som er gelatinøs (→ bevæger sig sammen med endolymfen)
• Ved hovedrotation (rotationsacceleration) bevæges a) den ossøse labyrint b) endolymfen • Endolymfen (og cupula) er langsommere om at ”komme i gang” (inerti) → cupula forskydes • Hårceller a la cochlea derformeres → depolarisering af afferente neuron. • Samme hastighed efter ca. 30 sek (uden andre stimuli vil hjernen ikke registrere acceleration) • De tre buegange på hver side er placeret i hvert sit plan → kan registrere bevægels e i alle
planer. • Parret (findes på både højre og venstre side) → større præcision. 3. Sacculus og utriculus: • Ligger foran buegangene, hhv. i vertikalplanet og horisontalplanet. • Hårceller sidder ansamlet som macula sacculi og utriculi. • Hårcellers stereocilier stikker ind i gelatinøse statokoniemembran. • Statokoniemembranen tynges af otolitter (calciumphosphat krystaller). • Bevæges ved lineære bevægelser af hovedet (lineær acceleration) • En enkelt hårcelle depolariseres ved en bestemt retning og hyperpolariseres ved modsatte. • Hårceller plantet således at alle retninger dækkes ind • Parret (findes på både højre og venstre side): større præcision. 4. Centrale ligevægtsbaner • Impulser løber til vestibulære kerner ved øvre medulla oblongata/nedre pons. • Forbindelser til:
a) Vestibulocerebellum (nuclei fastigii) b) Øjenmuskler (via fasciculus longitudinalis medialis) c) Formatio reticularis (til rygmarv via tractus reticulospinalis) d) Rygmarven (i tractus vestibulospinalis) e) Få projektioner til cortex (rumlig opfattelse?)
Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 20 af 31
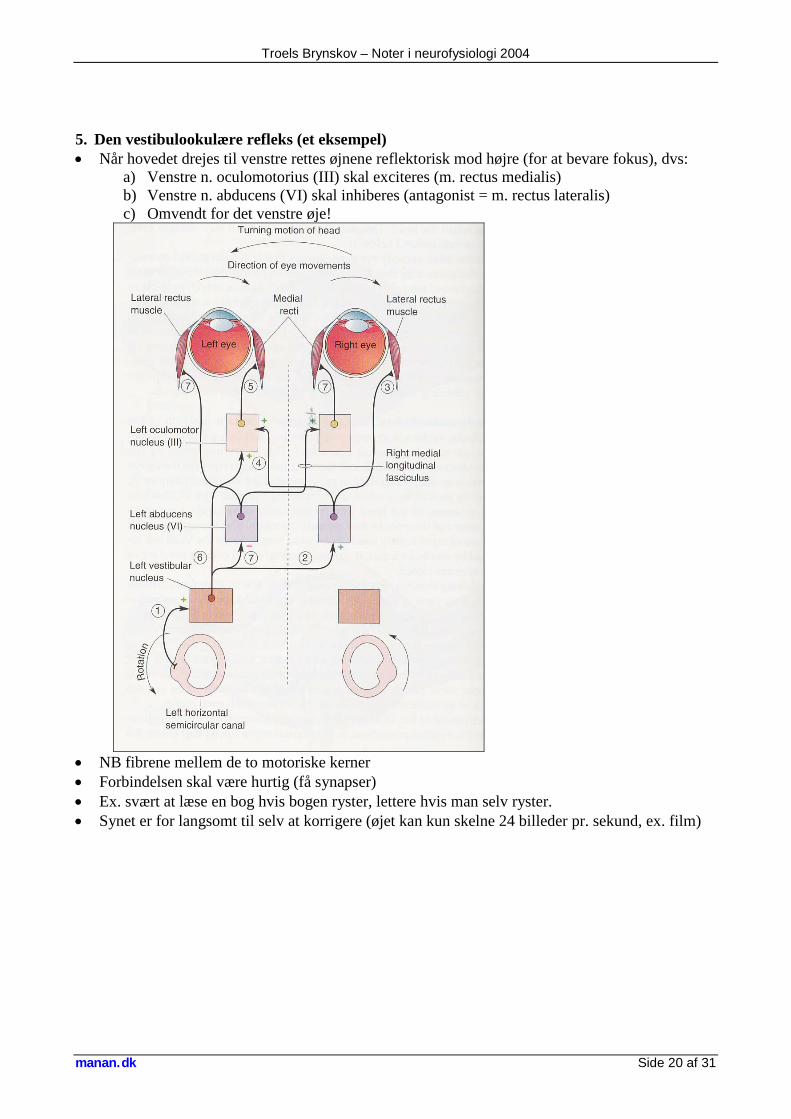
5. Den vestibulookulære refleks (et eksempel) • Når hovedet drejes til venstre rettes øjnene reflektorisk mod højre (for at bevare fokus), dvs:
a) Venstre n. oculomotorius (III) skal exciteres (m. rectus medialis) b) Venstre n. abducens (VI) skal inhiberes (antagonist = m. rectus lateralis) c) Omvendt for det venstre øje!
• NB fibrene mellem de to motoriske kerner • Forbindelsen skal være hurtig (få synapser) • Ex. svært at læse en bog hvis bogen ryster, lettere hvis man selv ryster. • Synet er for langsomt til selv at korrigere (øjet kan kun skelne 24 billeder pr. sekund, ex. film)

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 21 af 31
7. SPINALE MEKANISMER FOR MOTORISK FUNKTION Reflekser
Disposition: Stikord
1. α-motorneuronet kraft = antal rekrutterede enheder + frekvens 2. Den myotatiske refleks og γ-motorneuronet muskelten - Ia aff, α og γ efferent. 3. Reciprok inhibition agonist vs. antagonist 4. Den invers myotatiske refleks golgi seneten - Ib aff 5. Fleksorrefleksen smerte aff, fleks i standben + krydset ekstension 6. Gang spinale pacemakerceller og fleksorrefleks
Mange motoriske funktioner er styret af spinale mekanismer (jf. hovedløse høns) Det sensoriske og motoriske system er gensidigt afhængige, et sensori-motorisk system. 1. α-motorneuronet • ”The final common pathway” til en motorisk enhed i en muskel. • Rekrutteres i forskellig rækkefølge afhængigt af ønskede styrke. • Kraften reguleres altså ved:
a) Antallet af aktiverede α-motorneuroner (størrelserne af deres motoriske enheder) b) α –motorneuronernes fyringsfrekvens
2. Et α motorneuron modtager input fra: a) Refleksbuer på spinalt niveau b) Descenderende baner (se emne 8)
2. Den myotatiske refleks og γ-motorneuronet Den myotatiske refleks (statisk mekanisme): • Muskelten (= intrafusale muskelfibre. En type proprioceptorer) registrerer muskel-længde. • Ved muskelforlængning sendes impuls til forhornet via hurtige Ia afferenter: • α-motorneuron der således forkorter musklen kompensatorisk • En monosynaptisk refleks. • Denne mekanisme aktiveres ved patella-refleksen og kaldes derfor også strækrefleksen eller
senerefleksen. γ-motorneuronet (dynamisk mekanisme): • Kan spænde muskeltenen (vs. α der indirekte afslapper den) • Også forbundet med muskeltenens afferenter:
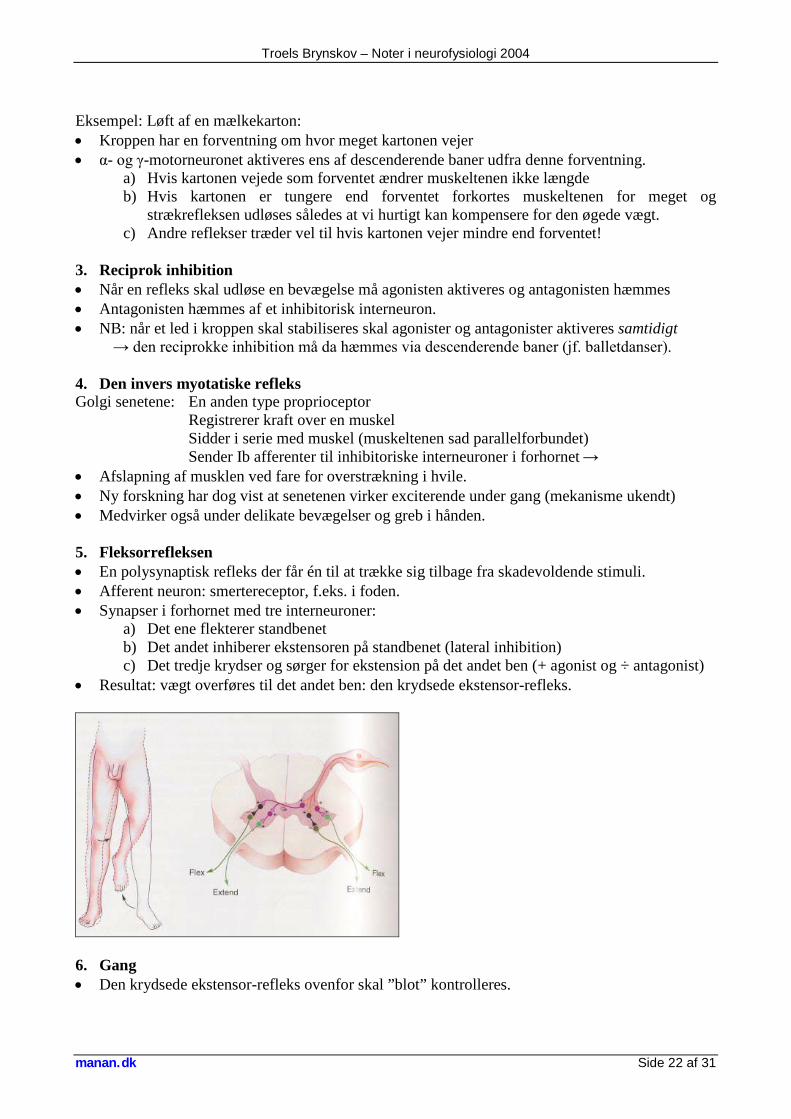
Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 22 af 31
Eksempel: Løft af en mælkekarton: • Kroppen har en forventning om hvor meget kartonen vejer • α- og γ-motorneuronet aktiveres ens af descenderende baner udfra denne forventning.
a) Hvis kartonen vejede som forventet ændrer muskeltenen ikke længde b) Hvis kartonen er tungere end forventet forkortes muskeltenen for meget og
strækrefleksen udløses således at vi hurtigt kan kompensere for den øgede vægt. c) Andre reflekser træder vel til hvis kartonen vejer mindre end forventet!
3. Reciprok inhibition • Når en refleks skal udløse en bevægelse må agonisten aktiveres og antagonisten hæmmes • Antagonisten hæmmes af et inhibitorisk interneuron. • NB: når et led i kroppen skal stabiliseres skal agonister og antagonister aktiveres samtidigt → den reciprokke inhibition må da hæmmes via descenderende baner (jf. balletdanser). 4. Den invers myotatiske refleks Golgi senetene: En anden type proprioceptor Registrerer kraft over en muskel Sidder i serie med muskel (muskeltenen sad parallelforbundet) Sender Ib afferenter til inhibitoriske interneuroner i forhornet → • Afslapning af musklen ved fare for overstrækning i hvile. • Ny forskning har dog vist at senetenen virker exciterende under gang (mekanisme ukendt) • Medvirker også under delikate bevægelser og greb i hånden. 5. Fleksorrefleksen • En polysynaptisk refleks der får én til at trække sig tilbage fra skadevoldende stimuli. • Afferent neuron: smertereceptor, f.eks. i foden. • Synapser i forhornet med tre interneuroner:
a) Det ene flekterer standbenet b) Det andet inhiberer ekstensoren på standbenet (lateral inhibition) c) Det tredje krydser og sørger for ekstension på det andet ben (+ agonist og ÷ antagonist)
• Resultat: vægt overføres til det andet ben: den krydsede ekstensor-refleks.
6. Gang • Den krydsede ekstensor-refleks ovenfor skal ”blot” kontrolleres.

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 23 af 31
• Der lader bla. til at være neuroner med pacemaker egenskaber (i rygmarven) der kan påvirke den krydsede ekstensor-refleks (kun påvist for katte og fisk!)
• Man kan tale om et spinalt gangcenter.

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 24 af 31
8. SUPRASPINALE MEKANISMER FOR MOTORISK FUNKTION Hvordan hjernen påvirker α-motorneuronet
Disposition: Stikord
1. Præfrontale cortex Klar, parat, start + PET scanning. 2. Basalganglierne Indirekte og direkte kredsløb. ”Bremse på area 6” 3. Primær motorisk cortex Homunculus, plasticitet 4. Pontocerebellum S1 – basalganglier – thalamus – M1. Bevægelsesprogram. 5. Descenderende baner forstreng (grovmotorik) og sidestreng (finmotorik)
1. Præfrontale cortex • Højere planlægning af hvad man vil foretage sig. • Aktivitet kan måles ca. ½ sek før man er bevidst om at ville udføre en bevægelse og 1 sek før
man rent faktisk udfører bevægelsen (dansk PET forsøg). • Arbejder med klar, parat, start:
a) Klar: Præmotoriske cortex b) Parat: Area 6 c) Start: Area 4
2. Basalganglierne • Et slags bremse-loop mellem præfrontale cortex og primær motorisk cortex (area 6) • Har altid en baggrundsfyring (baggrundsbremse) • To kredsløb:
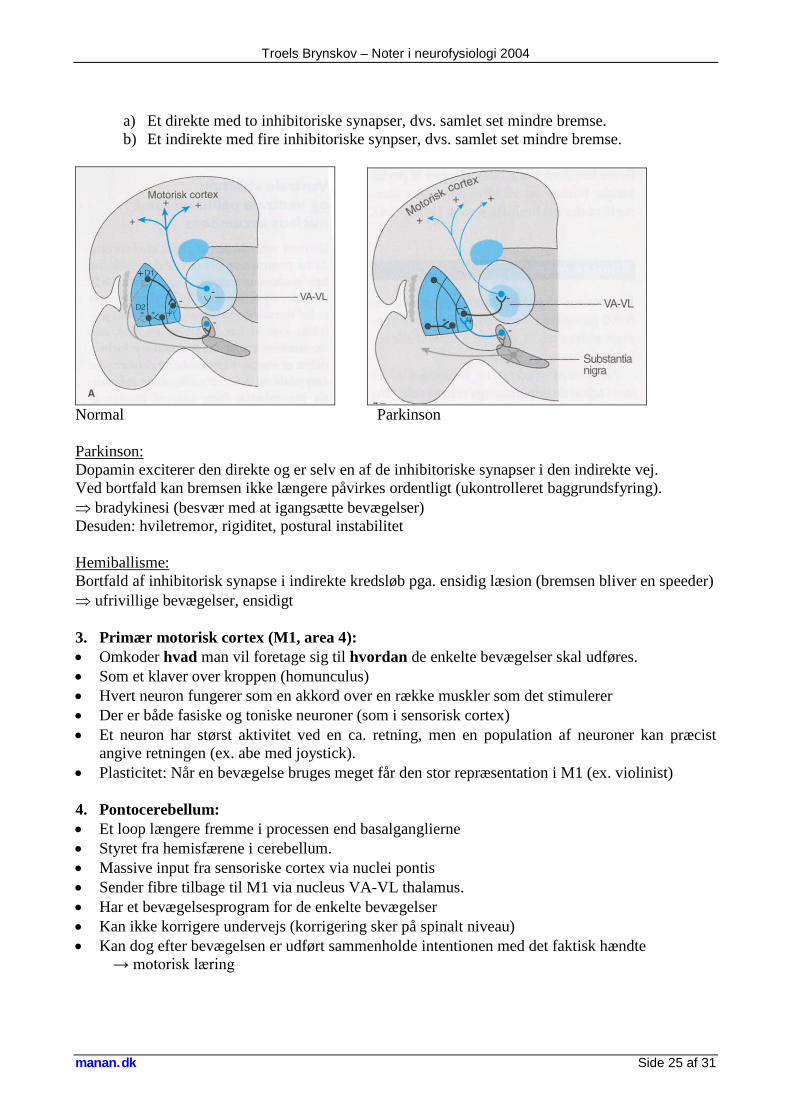
Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 25 af 31
a) Et direkte med to inhibitoriske synapser, dvs. samlet set mindre bremse. b) Et indirekte med fire inhibitoriske synpser, dvs. samlet set mindre bremse.
Normal Parkinson Parkinson: Dopamin exciterer den direkte og er selv en af de inhibitoriske synapser i den indirekte vej. Ved bortfald kan bremsen ikke længere påvirkes ordentligt (ukontrolleret baggrundsfyring). ⇒ bradykinesi (besvær med at igangsætte bevægelser) Desuden: hviletremor, rigiditet, postural instabilitet Hemiballisme: Bortfald af inhibitorisk synapse i indirekte kredsløb pga. ensidig læsion (bremsen bliver en speeder) ⇒ ufrivillige bevægelser, ensidigt 3. Primær motorisk cortex (M1, area 4): • Omkoder hvad man vil foretage sig til hvordan de enkelte bevægelser skal udføres. • Som et klaver over kroppen (homunculus) • Hvert neuron fungerer som en akkord over en række muskler som det stimulerer • Der er både fasiske og toniske neuroner (som i sensorisk cortex) • Et neuron har størst aktivitet ved en ca. retning, men en population af neuroner kan præcist
angive retningen (ex. abe med joystick). • Plasticitet: Når en bevægelse bruges meget får den stor repræsentation i M1 (ex. violinist) 4. Pontocerebellum: • Et loop længere fremme i processen end basalganglierne • Styret fra hemisfærene i cerebellum. • Massive input fra sensoriske cortex via nuclei pontis • Sender fibre tilbage til M1 via nucleus VA-VL thalamus. • Har et bevægelsesprogram for de enkelte bevægelser • Kan ikke korrigere undervejs (korrigering sker på spinalt niveau) • Kan dog efter bevægelsen er udført sammenholde intentionen med det faktisk hændte → motorisk læring

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 26 af 31
Klinik: Ved læsion af pontocerebellum ses: • Ataxi = ukoordinerede og upræcise bevægelser. Store udsving når pegefinger føres til næsen.
Rammer dog næsen! • Ses også ved alkoholpåvirkning. 5. Descenderende baner: • Sidestrengsbanerne styrer finmotorik
a) Den corticospinale bane (de fibre der krydser i decussatio pyramis) b) Den rubrospinale bane
• Forstrengsbanerne styrer grovmotorik a) Den reticulospinale bane (fastholder eller ophæver stand-reflekser og påvirker smerte) b) Den vestibulospinale bane (holder hovedet balanceret ift. kroppen) c) Den tectospinale bane (reagerer på ændrede lyde og syns-stimuli) d) Den corticospinale bane (de fibre der ikke krydser i decussatio pyramis)

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 27 af 31
9. HJERNENS RYTMER Biologiske rytmer
Disposition:
1. Elektroencephalogram, EEG 2. Epilepsi 3. Søvn 4. Døgnrytmer (circadiske rytmer)
Elektroencephalogram, EEG: • Måler synkrone (summerede) synaptiske potentialer i cortex. • Kun et yderst ringe bidrag fra propagerende axionspotentialer. • Enkelt synapses bidrag er ikke målbart gennem skalpen • Område på omkring 2 mm2 dækkes af hver elektrode • Amplitude: Øget synkronitet giver større amplitude • Frekvens: Øget synkronitet giver lavere frekvens (theta rytmer) • Største synkronitet: dyb søvn, mindste synkronitet: vågen/REM søvn (beta rytmer) • Bruges til:
a) At diagnosticere epilepsi b) At undersøge søvnrytmer/forstyrrelser
Epilepsi: ”Heterogen gruppe af sygdomme karakteriseret ved abnorm grad af synkronisering af større eller mindre grupper af neuroner” • Epileptiske anfaldsformer:
a) Generel krampe (hele cortex, begge hemisfærer) b) Fokal krampe (afgrænset område i cortex)
• EEG mønsteret er meget abnormt under (iktale) og mellem (interiktale) et epileptisk anfald. Søvn: EEG rytmer:
a alfa: 8-12 Hz beta: 12-30 Hz theta: 4-8 Hz alfa spindler og k-komplekser delta 1: 1-4 Hz delta 2: <1 Hz
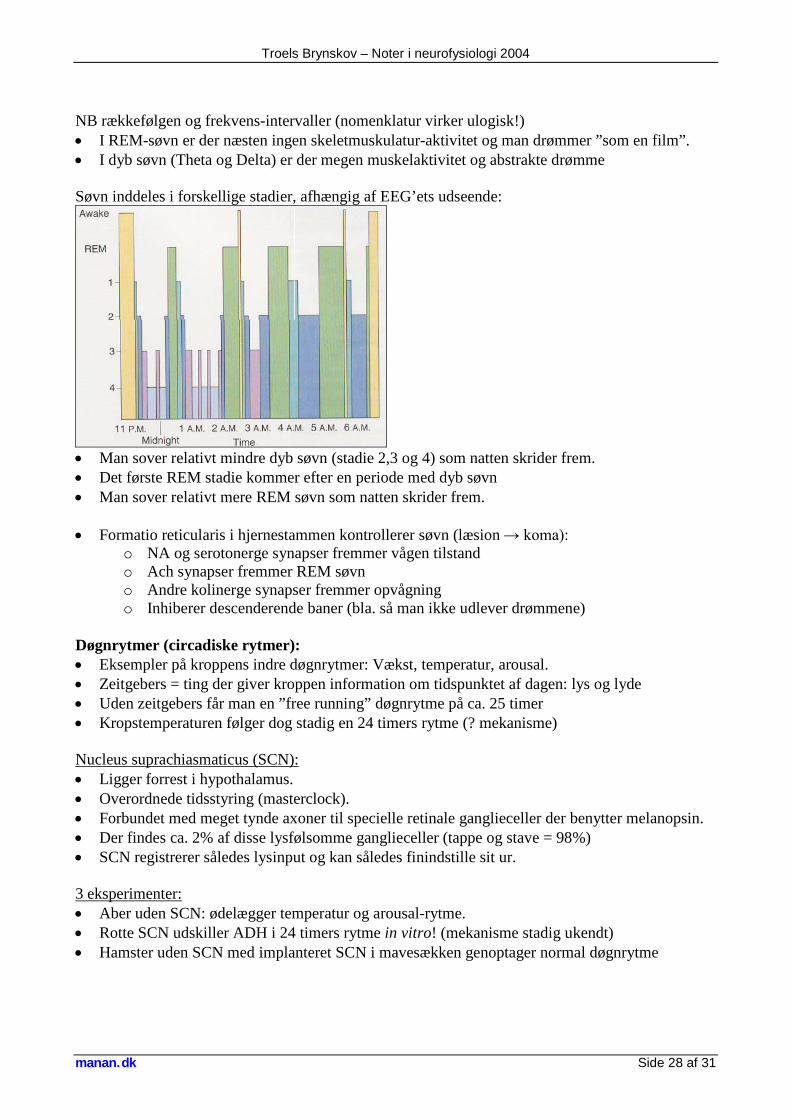
Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 28 af 31
NB rækkefølgen og frekvens-intervaller (nomenklatur virker ulogisk!) • I REM-søvn er der næsten ingen skeletmuskulatur-aktivitet og man drømmer ”som en film”. • I dyb søvn (Theta og Delta) er der megen muskelaktivitet og abstrakte drømme Søvn inddeles i forskellige stadier, afhængig af EEG’ets udseende:
• Man sover relativt mindre dyb søvn (stadie 2,3 og 4) som natten skrider frem. • Det første REM stadie kommer efter en periode med dyb søvn • Man sover relativt mere REM søvn som natten skrider frem. • Formatio reticularis i hjernestammen kontrollerer søvn (læsion → koma):
o NA og serotonerge synapser fremmer vågen tilstand o Ach synapser fremmer REM søvn o Andre kolinerge synapser fremmer opvågning o Inhiberer descenderende baner (bla. så man ikke udlever drømmene)
Døgnrytmer (circadiske rytmer): • Eksempler på kroppens indre døgnrytmer: Vækst, temperatur, arousal. • Zeitgebers = ting der giver kroppen information om tidspunktet af dagen: lys og lyde • Uden zeitgebers får man en ”free running” døgnrytme på ca. 25 timer • Kropstemperaturen følger dog stadig en 24 timers rytme (? mekanisme) Nucleus suprachiasmaticus (SCN): • Ligger forrest i hypothalamus. • Overordnede tidsstyring (masterclock). • Forbundet med meget tynde axoner til specielle retinale ganglieceller der benytter melanopsin. • Der findes ca. 2% af disse lysfølsomme ganglieceller (tappe og stave = 98%) • SCN registrerer således lysinput og kan således finindstille sit ur. 3 eksperimenter: • Aber uden SCN: ødelægger temperatur og arousal-rytme. • Rotte SCN udskiller ADH i 24 timers rytme in vitro! (mekanisme stadig ukendt) • Hamster uden SCN med implanteret SCN i mavesækken genoptager normal døgnrytme

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 29 af 31
10. HUKOMMELSE og lidt om indlæring
Disposition: 1. Lagring af information og forskellige typer hukommelse 2. Lokalisering af hukommelse 3. Molekylære mekanismer
• Hjernen fungerer på en anden måde end en computer harddisk. • Mere til helheder end detaljer (eksempelvis genkendelse af billeder og ansigter). • Plasticitet. • Meget stor kapacitet. Øverste grænse kendes ikke (hvis den findes) 1. Lagring af information og forskellige typer hukommelse Meget simplificeret model af lagringen og genkaldelse i hjernen: Amnesi: • Mistet hukommelse eller evne til indlæring. • Retrograd = mistet hukommelse fra før et traume, kan huske siden (genkendelse er ramt) • Anterograd = mistet dele af hukommelse efter et traume, men kan huske umiddelbart før.
(konsolidering er ramt) • Hvad der var lagret i korttidshukommelse forsvinder f.eks. ved milde traumer og ECT (ren
amnesi) Forskellig typer hukommelse: Deklarativ hukommelse: • Fakta og begivenheder: hvad, hvor og hvornår. • Hurtigt etableret, men labil • Sker bla. i temporallappen. Procedural hukommelse: • Bevægelsesfærdigheder; hvordan man gør ting. • langsomt etableret, men stabil • Sker bla. i cerebellum og striatum. Andre, mere specielle, typer hukommelse findes også.
a) ”Working memory” b) Korttidshukommelse
labil (flygtig)
langtidshukommelse stabil
Genkendelse
Konsolidering
20-30 min
Sanse-indtryk
Svækkes med alderen

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 30 af 31
2. Lokaliseringen af hukommelse Lashleys rottestudier (1920’erne): • Optrænede en rotte til at gennemføre en labyrint • Fjernede dernæst både forskellige dele af cortex og forskellige størrelser • Lashley talte derpå dens fejl i labyrinten. • Konkluderede at det var mængden af cortex fjernet der var det afgørende, ikke stedet. • Forkert at hukommelse er ligeligt fordelt, rigtigt at hukommelse er spredt i cortex Hebbs hypotese (1949): • En samling neuroner aktiveres af et stimulus. • Først aktiveres neuronerne og bliver aktive i kort tid (korttidshukommelse) • Hvis aktiveringen er ”kraftig nok” vil de danne reciprokke forbindelser (langtidshukommelse) • En senere aktivering af dele af disse neuroner vil genkalde hele det oprindelige stimulus • Stadig en populær model over hjernens måde at indlære, lagre og genkalde
• Kan sammenholdes med relational memory teorien, dvs. at hjernen lagrer al deklarativ
hukommelse lært på det samme tidspunkt – man kan senere genkalde den ene ting (ex. ”det man læste i bogen”) via de andre omstændigheder man kosoliderede det (”sidde i togkupeen”, ”naboen spillede høj musik”)
Milners studier af HM (1953): • HM havde svær epilepsi og fik fjernet begge temporallapper (!) • Indgrebet hjalp på epilepsien

Troels Brynskov – Noter i neurofysiologi 2004
manan.dk Side 31 af 31
• Men HM har mistet evnen til at skabe ny (kort- og langtids-) deklarativ hukommelse • = anterograd amnesi • Kan lære med sin procedurale hukommelse Oltons labyrint og working memory: • Stjerneformet labyrint med flere arme hvor rotten kan lære hvilke arme der aldrig er mad i. • Læsioner i hippocampus får rotten til at vælge arme hvor den allerede har været. • Den har således problemer med sin working memory • Den kan dog godt lære sig hvilke arme der aldrig er mad i (ok langtidshukommelse) • Kan også lære denne øvelse med lamper der angiver hvor der er mad (ok procedural
hukommelse) • Læsioner i striatum kan ikke finde ud af lampeversionen, men går aldrig to gange til den samme
gang. Lokalisering som den kendes i dag: • Hippocampus: working memory, spatial hukommelse. • Temporallappen: se HM • Cerebellum og striatum: procedural hukommelse • Diencephalon: konsolidering • Amygdala: emotionel hukommelse 3. Moleylære mekanismer for lagring (EJ KOMPLET!!!) Flere forskellige delvist isolerede landvindinger: Proteinsyntesehæmmer til rotter: • blabla • blabla CREB hos bananfluen: • Normal bananflue kan lære at undgå at flyve til det rør hvor den får stød. Glemmer dog igen. • Ved defekt CREB-gen kan den slet ikke lære det. • Ved overrekspression af CREB-gen glemmer den det aldrig. Stimulerede rotter: • Velstimulerede rotter danner flere synapser i deres cortex end ustimulerede rotter gør Slices af hippocampus (Bear s. 733): • LTP/D • CREB Stedceller i rotter, LTP: • blabla Rettelser, tilføjelser og/eller ideer til disse noter modtages gerne på [email protected]. Noten findes også på www.brynskov.com/troels/3sem





























