-
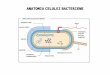
STRUCTURA CELULEI BACTERIENE Unitatea morfofuncional a
bacteriilor este celula procariot , un complex autoorganizat de
molecule organice care schimb energie cu mediul nconjurtor i este
capabil s -i regleze n mod autonom funciile vitale.
Curs 3
-
ComponenteObligatoriiNucleuCitoplasm MezozomiMembran
citoplasmatic Perete celularNeobligatoriiCapsul
FlageliFimbriispor
-
Structura celulei bacteriene
-
Nucleul bacterian = nucleoid,echivalent nuclearstructur primitiv
reprezint 2% din greutatea uscat a bacteriei. molecul circular de
ADN, organizat sub forma unui cromozom haploid care este n contact
direct cu citoplasma datorit lipsei membranei nuclearemolecula de
ADN dublu spiralat este la rndul ei suprahelicat n jurul unui miez
de ARN, dispoziie necesar funcionalit ii materialului nuclearsituat
n centrul celulei bacteriene i legat de membrana citoplasmatic prin
intermediul mezozomilor.
-
Funcia nucleului :depozitarea informaiei genetice necesar
autoreplic rii, organiz rii structurale i funcionale a celulei
bacteriene, deci a caracterelor ce definesc specia.n afar de ADN
cromozomial, la unele bacterii sunt prezente molecule circulare
mici, extracromozomiale de ADN care se numesc plasmide i care se
replic independent de cromozomul bacterian.
-
Citoplasmaun sistem coloidal alctuit din 80% ap, n care se gsete
o cantitate mare de molecule organice, ioni anorganici, enzime, ARN
(de transport, mesager i ribozomial), vacuole i incluzii. lipsit de
organitele celulare prezente la celulele eucariote cum sunt RE,
aparatul Golgi, mitocondriile, centrul celular, ergastoplasma
20.000 de ribozomi ce conin 80-90% din ARN-ul citoplasmatic
Ribozomii sunt structuri sferice cu diametru de aproximativ 180nm i
constanta de sedimentare de 70S. n absena ionilor de Mg2+ se desfac
n dou subuniti cu constanta de sedimentare de 50S i 30 S. Din punct
de vedere chimic sunt alctuii din 65-70% ARN i 30-35% proteine. O
parte din ribozomi se asociaz formnd polizomi (mai ales n timpul
sintezelor proteice), alii sunt liberi, iar o a treia categorie se
ataeaz mezozomilor sau membranei citoplasmatice.reprezint sediul
sintezelor proteice din celul Incluziile citoplasmatice: formaiuni
structurale inerte, temporare cu rol de rezerv energetic
-
Membrana citoplasmatic
-
o membran fin, elastic, lipsit de rezisten mecanic pare
trilaminat, fiind alctuit din dou straturi fosfolipidice dispuse cu
prile hidrofobe fa n fa se gsesc molecule proteice, situate fie la
nivelul unuia dintre cele dou straturi fosfolipidice, fie le
traverseaz fiind expuse la ambele fee ale membraneiStructura
membranei citoplasmatice
-
Functiile membranei citoplasmatice are permeabilitate selectiv,
ndeplinind funcia de barier osmotic ce regleaz schimburile celulei
bacteriene, n ambele sensuri cu mediul inconjurtorsecret enzime
hidrolitice ce se elibereaz n mediul nconjurtor unde scindeaz
substratul nutritiv n uniti absorbabile unele proteine legate de
membrana citoplasmatic joac rolul de chemoreceptori sediul
enzimelor lanului respirator i al fosforilrii oxidative, deci
centrul energogenezei celulare implicat n sinteza peretelui
celular, al polizaharidelor capsulare activ la creterea i
diviziunea celulei bacteriene, la formarea sporului bacterian
constituie o posibil int pentru aciunea unor chimioterapice ca, de
pild, polimixinele
-
Mezozomii se formeaz prin invaginarea membranei citoplasmatice
nu formeaz n citoplasm caviti inerte, ci deschise spre spaiul
periplasmic (spaiul dintre membrana citoplasmatic i peretele
celular) i sunt n contact direct cu materialul nuclearFunciile
mezozomilorparticip la replicarea cromozomului bacterian i
diviziunea celular,particip la reaciile de fosforilare oxidativ i
oxidoreducere, dar n msur mai mic dect membrana celular,sediul unor
enzime hidrolitice care ndeplinesc rolul enzimelor lizozomale de la
celulele eucariote,sinteza i secreia unor exoenzime, ca de exemplu
penicilinaza sau cefalosporinaza,sinteza peretelui celular i
formarea sporului bacterian
-
Peretele celular
structur rigid specific bacteriilor, care nconjoar membrana
citoplasmatic strat bazal, asemntor la toate bacteriile un strat al
structurilor superficiale, foarte difereniat, n funcie de care
bacteriile manifest caractere tinctoriale diferite: bacterii gram
pozitivegram- negative i acido-alcoolorezistente
-
Peretele celular- stratul bazal
-
Structura stratului bazal. Peptidoglicanul
- structura chimic responsabil de rigiditatea peretelui celular-
asigur forma i rezistena mecanic a bacteriei- const dintr-un
schelet, format din molecule lungi paralele polizaharidice de
N-acetil-glucozamin i acid N-acetil-muramic. - moleculele de acid
N-acetil-muramic din lanurile vecine sunt legate ntre ele prin puni
polipeptidice, transversale, a cror structur difer la bacteriile
gram- pozitive de cele gram-negative i chiar de la specie la
specie. - de fiecare molecul de acid N-acetil-muramic se leag un
tetrapeptid ce conine D i L aminoacizi.
-
Structura stratului bazal. 2.La bacteriile gram +pozitive dou
tetrapeptide de pe 2 lanuri paralele nvecinate sunt legate
printr-un pentapeptid de glicina. La bacteriile gram - negative, -
tetrapeptidele se leag ntre ele printr-o simpl legtur peptidic.
Menionam c acidul diaminopimelic i acidul N-acetil-muramic sunt
componente care nu au fost gsite n natur dect la bacterii.
-
Structura stratului superficial
-
Structura stratului structurilor specialeDifera in functie de
tipul bacteriei :gram-pozitiva, gram-negativa
acido-alcoolorezistenta.La bacteriile gram +
pozitivepeptidoglicanul reprezint 50-90% din greutatea uscat a
peretelui celulargrosime de 15-30 nm200 de lanuri paralele de
murein.
-
. Deosebim 3 tipuri de structuri speciale: gram-pozitiv,
gram-negativ i acido-alcoolorezistent.La bacteriile gram+pozitive,
peptidoglicanul reprezint 50-90% din greutatea uscat a peretelui
celular, are o grosime de 15-30 nm i conine pn la 200 de lanuri
paralele de murein.Stratul structurilor speciale este redus i
alctuit din polimeri hidrosolubili care sunt acizii theicoici:
acidul ribitoltheicoic i gliceroltheicoic. Acetia pot ptrunde pn la
membrana citoplasmatic legndu-se covalent de aceast - acizii
theicoici de membran sau numai pn la perete - acizi theicoici de
perete. Ei reprezint antigenele de supafa ale bacteriilor
gram-pozitive. Peretele celular al bacteriilor gram-pozitive este
sensibil la aciunea lizozimului care rupe legturile dintre acidul
N-acetil muramic i N-acetilglucozamin i a penicilinei care nhib
sinteza peptidoglicanului.
redus i alctuit din acizi theicoici: acidul ribitoltheicoic
acidul gliceroltheicoic. Acetia pot ptrunde pn la membrana
citoplasmatic legndu-se covalent de aceast - acizii theicoici de
membran sau numai pn la perete - acizi theicoici de perete. Ei
reprezint antigenele de supafa ale bacteriilor gram-pozitive.
Stratul structurilor speciale
-
Peretele celular-2-Peretele celular al bacteriilor gram-pozitive
este sensibil la aciunea lizozimului care rupe legturile dintre
acidul N-acetil muramic i N-acetilglucozamin i a penicilinei care
nhib sinteza peptidoglicanului.
-
La bacteriile gram-negative,peptidoglicanul are o grosime de
4-5nm, reprezentnd numai 10% din greutatea uscat a bacteriei. Gram
negativii prezinta 3 structuri :membran extern, Lipoproteine
lipopolizaharidul de perete.
-
Membrana externaMembrana extern este format dintr-un strat dublu
fosfolipoproteic ce cuprinde o cantitate foarte mare de molecule
proteice. Proteinele nespecifice ale membranei externe cu funcie de
porine, cum sunt trimerii OmpF i OmpC, delimiteaz nite canale
umplute cu ap prin intermediul crora proteinele pot difuza n spaiul
periplasmic -substane hidrosolubile.
-
Proteinele specifice Proteinele specifice au funcii de transport
pentru anumite substane, unele dintre ele fiind i receptori pentru
bacteriofagi. Proteinele de suprafa pot avea la unele bacterii rol
n patogenitate, ca de exemplu cele 7 proteine de suprafa al
shigellelor care le asigur invazivitatea.
-
LPZ sau endotoxinaDeasupra membranei externe a bacililor
gram-negativi se afl lipopolizaharidul de perete (LPZ) sau
endotoxina bacililor gram-negativi. Aceasta este alctuit
din:lipidul A care are o structur particular, fiind format din
uniti dizaharidice de glucozamin, legate de beta-hidroxiacizi grai
cu 10-16 atomi de carbon fixai direct de membrana extern,miezul sau
core, numit i antigen R, comun tuturor bacteriilor gram -.uniti
monozaharidice repetitive (15-40) care sunt specifice de specie i
tip i constituie antigenul O al bacteriilor gram-negative.
-
LPZ -2. i Spatiul periplasmicLPZ este o toxin termolabil, care
se elibereaz n mediul nconjurtor de ctre bacteriile gram-negative
numai dup liza lor i foarte reactiv n organismul gazd. Astfel,
lipidul A produce febr, activeaz mecanismele aprrii antiinfecioase
i n exces produce ocul endotoxic cu evoluie grav, chiar
fatal.Spaiul periplasmic. Sistemul structural dublu al peretelui
celular la bacteriile gram-negative creeaz un compartiment ce se
ntinde de la membrana celular pn la membrana extern, numit spaiu
periplasmic. El conine peptidoglicanul i un gel care favorizeaz
nutriia bacteriei prin coninutul n enzime degradative ca, de pild,
fosfataze, nucleaze, proteaze etc. Tot aici sunt prezente enzimele
de inactivare ale unor antibiotice cum sunt beta-lactamazele i
cefalosporinazele.
-
Bacteriile acido-alcoolo-rezistente Bacteriile
acido-alcoolorezistente de interes medical sunt bacilul tuberculos
i bacilul leprei a cror perete celular se aseamn cu cel al
bacteriilor gram-pozitive. Structurile speciale conin acid micolic
i o cear ce confer acestor bacterii caractere tinctoriale deosebite
i rezisten crescut la factorii de mediu. Astfel dac, dup o nclzire
de scurt durat ce topete cerurile, un colorant ptrunde n celula
bacterian, decolorarea acesteia sub aciunea acizilor sau alcoolilor
nu se mai produce ca la alte bacterii. Acest caracter se numete
acido-alcoolo-rezisten. Bacteriile acido-acoolo-rezistente se
coloreaz foarte slab cu coloraia gram, evidenierea lor fcndu-se la
cald prin tehnica Ziehl-Neelsen.
-
Funciile peretelui celularasigur forma, rezistena mecanic i
osmotic a bacteriei;asigur protecia membranei citoplasmatice fa de
presiunea intern a bacteriei (5-6 atm la E. Coli)regleaz traficul
molecular de perete n ambele sensuri, deci schimbul de substane
dintre bacterie i mediul nconjurtor, fiind permeabil pentru
molecule cu o GM mai mic de 10.000 daltoni i cu un diametru mai mic
de 1nm;
-
stocheaz unele enzime n spaiul periplasmic la bacteriile
gram-negative ce vor fi eliberate dup necesiti, spre deosebire de
bacteriile gram-pozitive care i elimin enzimele direct n mediul
extern;prezint receptori pentru bacteriofagi;este sediul
antigenelor de suprafa, fiind deci implicat n rspunsul imun al
macroorganismului;este sediul unor factori de patogenitateare rol n
diviziunea bacterian i n procesul de sporulare.
-
Componente bacteriene neobligatorii Capsulanveli compact, intim
legat de celula bacterian, cu o lime de 0,2de natur polizaharidic
(Streptococcus pneumoniae, Klebsiella etc.) cu excepia capsulei
bacilului crbunos care este polipeptidic.Variantele necapsulate ale
acelorai specii sunt nepatogeneCapsula este o structur cu proprieti
antigenice specifice (antigenele K) care permit diferenierea unor
serotipuri n cadrul speciei.
-
Microcapsulaeste o structur discret cu o grosime sub 0,2m care
nu se evideniaz la microscopul optic, ci numai prin metode
imunologice sau electronomicroscopice (Neisseria gonorrhoeae). Ea
constituie un factor de virulen Stratul mucosstrat amorf i vscos ce
nvelete bacteria. El este format din lanuri lungi de polizaharide,
cum sunt levanii i dextranii, cu rol major n adezivitatea
bacteriilor de suprafee.
-
Funciile capsulei:este un factor de aderen i colonizare a
bacteriilor pe suprafee;protejeaz bacteriile de diferii ageni
antibacterieni din mediu cum sunt: bacteriofagii, colicinele,
complementul, lizozimul sau alte enzime bacteriolitice;protejeaz
bacteriile de aciunea fagocitelor, fiind deci un factor de
virulen;reprezint sediul antigenelor capsulare, importante n
identificarea acestor bacterii.
-
Flagelii sau cilii
-
Pilii bacterieniPilii bacterienipe suprafaa bacteriilor
gram-negative apendici filamentoi, rigizi, mai scuri dect
flageliicu dispoziie n general peritriche se evideniaz prin
microscopie electronic. polimeri proteici de pilin, dispui
helicoidal ntr-o structur tubularproprieti antigenice.
-
Din punct de vedere funcional, :sex pili - codificai de
formaiunile genetice extracromozomiale (plasmide) implicai n
transferul de material genetic ntre bacterii. Sunt prezeni mai ales
la bacteriile gram-negative (Enterobacteriaceae, Pseudomonas), pili
comuni sau pili de aderen (fimbrii), n numr mare, codificai
cromozomial, cu rol in fixarea ferm de mediul de cultur sau de
celulele organismului. Constituie un factor important de virulen
(de exemplu la gonococ),au i proprieti antifagocitare.ambele
categorii de pili prezint antigene specifice piliare i pot fi
receptori pentru bacteriofagi.
-
Sporul bacterianSPORUL- O structura de rezistenta si
patogenitate a bacteriilor. Bacteriile se sporuleaza atunci cand
conditiile de mediu ( temperatura, uscaciune, lipsa de oxigen, etc)
devin neprielnice pentru viata si inmultirea bacteriilor.
Sporularea creste si rezistenta la Antibiotice.
-
Structura sporului core sau protoplastul sporal, format din
materialul nuclear inconjurat de membrana citoplasmatic. Aici este
depozitat DNA i o substan care are rol n rezistena sporului la
cldur - dipicolinatul de Ca, care nu s-a mai evideniat sanicieri n
natur, cortexul intern sau peretele sporal format din peptidoglican
i care este peretele celular primordial, membrana extern sau
cortexul sporal, care este stratul cel mai gros al sporului i care
conine i el un peptidoglican, cu o structur particular, foarte
sensibil la lizozim. tunica proteic este format din proteine
chitinoase cu numeroase legturi disulfitice. Ea este impermeabil,
fiind responsabil de rezistena sporilor la unele dezinfectante,
exosporiumul este prezent numai la unii spori i conine lipoproteine
i zaharide.x
-
Structura sporuluiCoreProtoplastul sporalM.citoplasmatic
DNADipicolinat de CaCortex internPG=Perete celular
primordialMembranaexternExosporiumLP +ZTunica proteica
-
Poziia sporului
-
SporulareaSporularea. Este un proces ce se declaneaz n condiii
nefavorabile de via, cnd bacteriile sunt lipsite de surse
nutritive. Ea presupune formarea unor structuri noi i dispariia
altor structuri caracteristice formei vegetative ale bacteriei.Din
punct de vedere morfologic, sporularea ncepe prin condensarea la un
capt al celulei a materialului nuclear. Acesta va fi separat de un
sept ce provine din invaginarea membranei citoplasmatice.
-
Sporularea .2.Septul crete, se desprinde de membrana
citoplasmatic i formeaz n jurul materialului nuclear o membran dubl
a crei fee care se privesc sunt cele ce sintetizeaz peretele
bacterian. Acesta este stadiul de prespor.
-
Sporularea .3.Cele dou lamele ale dublei membrane presporale vor
sintetiza nveliurile sporale:
1. peretele sporal i cortexul situate ntre cele dou lamele ale
membranei presporale i 2. o tunic proteic i
3. exosporiumul - n afara membranei presporale.
-
Sporii
-
n condiii de mediu favorabile, sporul germineaz i va da natere
unei bacterii vegetative identice cu aceea n care s-a format.
Germinarea are loc n 3 etape:1. Activarea sporului. Se produce cnd
acesta ntlnete condiii favorabile de via, dar i un factor care s
lezeze nveliul sporal (mecanic sau chimic ca, de exemplu,
lizozimul, compui cu grupri sulfhidrilice libere etc.).Germinarea
sporului - detalii
-
2.Iniierea formarii sporului este declanat de un mediu nutritiv
bogat. Pentru unii spori triggerul este L-alanina, iar pentru
altele adenozina. Acesta duce la activarea unei enzime autolitice
care degradeaz cortexul sporal. Are loc absorbia de ap, eliberarea
dipicolinatului de calciu i degradarea unor compui sporali.
3.Dezvoltarea sporului. Protoplastul sporal se transform n
bacterie vegetativ, care trece printr-o perioad metabolic activ de
refacere a constituenilor celulari normali i a echipamentului
enzimatic complet
-
Germinarea sporului
**Situat ntre materialul nuclear i faa intern a membranei
citoplasmatice, citoplasma este un sistem coloidal alctuit din 80%
ap, n care se gsete o cantitate mare de molecule organice, ioni
anorganici, enzime, ARN (de transport, mesager i ribozomial),
vacuole i incluzii. Este lipsit de organitele celulare prezente la
celulele eucariote cum sunt reticulul endoplasmatic, aparatul
Golgi, mitocondriile, centrul celular, ergastoplasma.La microscopul
optic citoplasma apare omogen, amorf, iar la cel electronic fin
granulat datorit unui numr mare de ribozomi. Se apreciaz c o celul
bacterian are 20.000 de ribozomi ce conin 80-90% din ARN-ul
citoplasmatic.Ribozomii sunt structuri sferice cu diametru de
aproximativ 180nm i constanta de sedimentare de 70S. n absena
ionilor de Mg2+ se desfac n dou subuniti cu constanta de
sedimentare de 50S i 30 S. Din punct de vedere chimic sunt alctuii
din 65-70% ARN i 30-35% proteine. O parte din ribozomi se asociaz
formnd polizomi (mai ales n timpul sintezelor proteice), alii sunt
liberi, iar o a treia categorie se ataeaz mezozomilor sau membranei
citoplasmatice. Ribozomii reprezint sediul sintezelor proteice din
celul.Incluziile citoplasmatice, descrise la unele specii
bacteriene, sunt formaiuni structurale inerte, temporare, de
diferite dimensiuni, variind n funcie de specia bacterian i
condiiile de mediu. Compoziia lor chimic este diferit, ele putnd fi
de glicogen sau amidon (la unii bacilii aerobi sporulai), lipide
(la genul Bacillus), polimetafosfai (sau incluziile de volutin
descrise mai nti la Spirillum volutans i apoi de Babe i Ernst la
bacilii difterici) etc. Incluziile sunt structuri legate de
activitatea metabolic a celulei bacteriene i reprezint un material
de rezerv care poate fi folosit ca surs de energie.**Structura
stratului bazal. Peptidoglicanul- structura chimic responsabil de
rigiditatea peretelui celular- asigur forma i rezistena mecanic a
bacterieil- const dintr-un schelet, format din molecule lungi
paralele polizaharidice de N-acetil-glucozamin i acid
N-acetil-muramic. m- moleculele de acid N-acetil-muramic din
lanurile vecine sunt legate ntre ele prin puni polipeptidice,
transversale, a cror structur difer la bacteriile gram- pozitive de
cele gram-negative i chiar de la specie la specie. - de fiecare
molecul de acid N-acetil-muramic se leag un tetrapeptid ce conine D
i L aminoacizi.La bacteriile gram - pozitive dou tetrapeptide de pe
2 lanuri paralele nvecinate sunt legate printr-un pentapeptid de
glicina. La bacteriile gram - negative, - tetrapeptidele se leag
ntre ele printr-o simpl legtur peptidic. Menionm c acidul
diaminopimelic i acidul N-acetil-muramic sunt componente care nu au
fost gsite n natur dect la bacterii.
***Structura stratului structurilor speciale. Deosebim 3 tipuri
de structuri speciale: gram-pozitiv, gram-negativ i
acido-alcoolorezistent.La bacteriile gram - pozitive,
peptidoglicanul reprezint 50-90% din greutatea uscat a peretelui
celular, are o grosime de 15-30 nm i conine pn la 200 de lanuri
paralele de murein.Stratul structurilor speciale este redus i
alctuit din polimeri hidrosolubili care sunt acizii theicoici:
acidul ribitoltheicoic i gliceroltheicoic. Acetia pot ptrunde pn la
membrana citoplasmatic legndu-se covalent de aceast - acizii
theicoici de membran sau numai pn la perete - acizi theicoici de
perete. Ei reprezint antigenele de supafa ale bacteriilor
gram-pozitive. Peretele celular al bacteriilor gram-pozitive este
sensibil la aciunea lizozimului care rupe legturile dintre acidul
N-acetil muramic i N-acetilglucozamin i a penicilinei care nhib
sinteza peptidoglicanului. *La bacteriile gram-negative,
peptidoglicanul are o grosime de 4-5nm, reprezentnd numai 10% din
greutatea uscat a bacteriei. Stratul superficial este ns mult mai
complex dect la bacteriile gram - pozitive fiind alctuit dintr-o
membran extern, lipoproteine i lipopolizaharidul de perete.Membrana
extern este format dintr-un strat dublu fosfolipoproteic ce
cuprinde o cantitate foarte mare de molecule proteice. Proteinele
nespecifice ale membranei externe cu funcie de porine, cum sunt
trimerii OmpF i OmpC, delimiteaz nite canale umplute cu ap prin
intermediul crora pot difuza n spaiul periplasmic substane
hidrosolubile.*Proteinele specifice au funcii de transport pentru
anumite substane, unele dintre ele fiind i receptori pentru
bacteriofagi. Proteinele de suprafa pot avea la unele bacterii rol
n patogenitate, ca de exemplu cele 7 proteine de suprafa al
shigellelor care le asigur invazivitatea.Deasupra membranei externe
a bacililor gram-negativi se afl lipopolizaharidul de perete (LPZ)
sau endotoxina bacililor gram-negativi. Aceasta este alctuit
din:lipidul A care are o structur particular, fiind format din
uniti dizaharidice de glucozamin, legate de beta-hidroxiacizi grai
cu 10-16 atomi de carbon fixai direct de membrana extern,miezul sau
core, numit i antigen R, comun tuturor bacteriilor gram -.uniti
monozaharidice repetitive (15-40) care sunt specifice de specie i
tip i constituie antigenul O al bacteriilor gram-negative.LPZ este
o toxin termolabil, care se elibereaz n mediul nconjurtor de ctre
bacteriile gram-negative numai dup liza lor i foarte reactiv n
organismul gazd. Astfel, lipidul A produce febr, activeaz
mecanismele aprrii antiinfecioase i n exces produce ocul endotoxic
cu evoluie grav, chiar fatal.Spaiul periplasmic. Sistemul
structural dublu al peretelui celular la bacteriile gram-negative
creeaz un compartiment ce se ntinde de la membrana celular pn la
membrana extern, numit spaiu periplasmic. El conine peptidoglicanul
i un gel care favorizeaz nutriia bacteriei prin coninutul n enzime
degradative ca, de pild, fosfataze, nucleaze, proteaze etc. Tot
aici sunt prezente enzimele de inactivare ale unor antibiotice cum
sunt beta-lactamazele i cefalosporinazele.Bacteriile
acido-alcoolorezistente de interes medical sunt bacilul tuberculos
i bacilul leprei a cror perete celular se aseamn cu cel al
bacteriilor gram-pozitive. Structurile speciale conin acid micolic
i o cear ce confer acestor bacterii caractere tinctoriale deosebite
i rezisten crescut la factorii de mediu. Astfel dac, dup o nclzire
de scurt durat ce topete cerurile, un colorant ptrunde n celula
bacterian, decolorarea acesteia sub aciunea acizilor sau alcoolilor
nu se mai produce ca la alte bacterii. Acest caracter se numete
acido-alcoolo-rezisten. Bacteriile acido-acoolo-rezistente se
coloreaz foarte slab cu coloraia gram, evidenierea lor fcndu-se la
cald prin tehnica Ziehl-Neelsen.
*Proteinele specifice au funcii de transport pentru anumite
substane, unele dintre ele fiind i receptori pentru bacteriofagi.
Proteinele de suprafa pot avea la unele bacterii rol n
patogenitate, ca de exemplu cele 7 proteine de suprafa al
shigellelor care le asigur invazivitatea.Deasupra membranei externe
a bacililor gram-negativi se afl lipopolizaharidul de perete (LPZ)
sau endotoxina bacililor gram-negativi. Aceasta este alctuit
din:lipidul A care are o structur particular, fiind format din
uniti dizaharidice de glucozamin, legate de beta-hidroxiacizi grai
cu 10-16 atomi de carbon fixai direct de membrana extern,miezul sau
core, numit i antigen R, comun tuturor bacteriilor gram -.uniti
monozaharidice repetitive (15-40) care sunt specifice de specie i
tip i constituie antigenul O al bacteriilor gram-negative.LPZ este
o toxin termolabil, care se elibereaz n mediul nconjurtor de ctre
bacteriile gram-negative numai dup liza lor i foarte reactiv n
organismul gazd. Astfel, lipidul A produce febr, activeaz
mecanismele aprrii antiinfecioase i n exces produce ocul endotoxic
cu evoluie grav, chiar fatal.Spaiul periplasmic. Sistemul
structural dublu al peretelui celular la bacteriile gram-negative
creeaz un compartiment ce se ntinde de la membrana celular pn la
membrana extern, numit spaiu periplasmic. El conine peptidoglicanul
i un gel care favorizeaz nutriia bacteriei prin coninutul n enzime
degradative ca, de pild, fosfataze, nucleaze, proteaze etc. Tot
aici sunt prezente enzimele de inactivare ale unor antibiotice cum
sunt beta-lactamazele i cefalosporinazele.Bacteriile
acido-alcoolorezistente de interes medical sunt bacilul tuberculos
i bacilul leprei a cror perete celular se aseamn cu cel al
bacteriilor gram-pozitive. Structurile speciale conin acid micolic
i o cear ce confer acestor bacterii caractere tinctoriale deosebite
i rezisten crescut la factorii de mediu. Astfel dac, dup o nclzire
de scurt durat ce topete cerurile, un colorant ptrunde n celula
bacterian, decolorarea acesteia sub aciunea acizilor sau alcoolilor
nu se mai produce ca la alte bacterii. Acest caracter se numete
acido-alcoolo-rezisten. Bacteriile acido-acoolo-rezistente se
coloreaz foarte slab cu coloraia gram, evidenierea lor fcndu-se la
cald prin tehnica Ziehl-Neelsen.
*Cilii sau flagelii bacterieni apendici filamentoi cu originea n
citoplasma bacterian, i servesc ca organe de locomoie. Ei sunt
prezeni mai ales la bacili (Enterobacteriaceae) dar i la unii coci
(enterococ). Cilii sunt structuri helicoidale, cu dimensiunea n
general mai mare dect cea a celulei bacteriene creia i aparin
(2-15/2-20 nm) i se evideniaz microscopic prin coloraii speciale.
dispoziia i numrul cililor sunt caracteristice speciei. S-au
descris bacterii atriche (fr cili), monotriche - cu un cil polar
(Vibro cholerae), lofotriche - cu un smoc de cili situat la unul
din polii bacteriei - (Pseudomonas fluorescens), amfitriche - cu
cilii situai la ambii poli ai bacteriei (la genul Spirillum) i
peritriche - cu cilii dispui pe ntreaga suprafa a bacteriei
(Salmonella, E. Coli, Proteus etc.). structura cililor este
tubular, ei fiind formai dintr-o protein contractil, flagelina, sub
form polimerizat. Cilul se ataeaz de bacterie printr-un corpuscul
bazal i un crlig de articulaie aflate n citoplasma bacteriei.
Corpusculul bazal se inser prin crligul de articulaie de membrana
citoplasmatic i peretele celular. mobilitatea i direcia micrii
cililor pot fi influenate de concentraia unor substane din mediu
(chimiotactism pozitiv sau negativ) i de temperatur. Ei sunt sediul
antigenelor flagelare (H), importante n identificarea bacteriilor.
cilii bacterieni nu se observ dect pe preparate colorate special. n
practica de rutin nu se evideniaz flagelii, ci mobilitatea
bacteriiloro fie prin examinarea lor pe un preparat nativ ntre lam
i lamel n care se urmresc micrile bacteriilor, o prin nsmnarea
tulpinii pe un mediu semisolid prin neparea n profunzime a
mediului. Dac bacteria crete nu numai pe traseul nsmnrii i difuzeaz
n mediu, ea este mobil.Pilii bacterieni pe suprafaa bacteriilor
speciile gram-negative apendici filamentoi, rigizi, mai scuri dect
flagelii cu dispoziie n general peritriche se evideniaz prin
microscopie electronic. polimeri proteici de pilin, dispui
helicoidal ntr-o structur tubular proprieti antigenice. Din punct
de vedere funcional, pilii se mpart n:sex pili - codificai de
formaiunile genetice extracromozomiale (plasmide) implicai n
transferul de material genetic ntre bacterii. Sunt prezeni mai ales
la bacteriile gram-negative (Enterobacteriaceae, Pseudomonas), pili
comuni sau pili de aderen (fimbrii), n numr mare, codificai
cromozomial, la fixarea ferm de mediul de cultur sau de celulele pe
care se afl. Ei constituie, deci, un factor important de virulen
(de exemplu la gonococ). n afar de aderen, pilii comuni mai au i
proprieti antifagocitare.ambele categorii de pili prezint antigene
specifice piliare i pot fi receptori pentru bacteriofagi. *Poziia
sporului bacterian constituie un caracter taxonomic. Poate fi
central, ca la bacilul crbunos (Bacillus anthracis), subterminal,
ca la B.cereus, sau terminal, ca bacilul tetanic (Cl.tetani).
*Sporularea. Este un proces ce se declaneaz n condiii
nefavorabile de via, cnd bacteriile sunt lipsite de surse
nutritive. Ea presupune formarea unor structuri noi i dispariia
altor structuri caracteristice formei vegetative ale bacteriei.Din
punct de vedere morfologic, sporularea ncepe prin condensarea la un
capt al celulei a materialului nuclear. Acesta va fi separat de un
sept ce provine din invaginarea membranei citoplasmatice. *Septul
crete, se desprinde de membrana citoplasmatic i formeaz n jurul
materialului nuclear o membran dubl a crei fee care se privesc sunt
cele ce sintetizeaz peretele bacterian. Acesta este stadiul de
prespor.Cele dou lamele ale membranei presporale vor sintetiza
nveliurile sporale: peretele sporal i cortexul situate ntre cele
dou lamele ale membranei presporale i o tunic proteic i
exosporiumul n afara membranei presporale.Structura sporului variaz
de la specie la specie, dar organizarea general se aseamn. De la
interior spre exterior, sporul este format din: core sau
protoplastul sporal, format din materialul nuclear inconjurat de
membrana citoplasmatic. Aici este depozitat DNA i o substan care
are rol n rezistena sporului la cldur - dipicolinatul de Ca, care
nu s-a mai evideniat nicieri n natur, cortexul intern sau peretele
sporal format din peptidoglican i care este peretele celular
primordial, membrana extern sau cortexul sporal, care este stratul
cel mai gros al sporului i care conine i el un peptidoglican, cu o
structur particular, foarte sensibil la lizozim. Autoliza acestui
strat este momentul cheie n transformarea sporului n forma
vegetativ, tunica proteic este format din proteine chitinoase cu
numeroase legturi disulfitice. Ea este impermeabil, fiind
responsabil de rezistena sporilor la unele dezinfectante,
exosporiumul este prezent numai la unii spori i conine lipoproteine
i zaharide.x*Germinarea sporului. n condiii de mediu favorabile,
sporul germineaz i va da natere unei bacterii vegetative identice
cu aceea n care s-a format. Germinarea are loc n 3 etape:Activarea
sporului. Se produce cnd acesta ntlnete condiii favorabile de via,
dar i un factor care s lezeze nveliul sporal (mecanic sau chimic
ca, de exemplu, lizozimul, compui cu grupri sulfhidrilice libere
etc.).Iniierea este declanat de un mediu nutritiv bogat. Pentru
unii spori triggerul este L-alanina, iar pentru altele adenozina.
Acesta duce la activarea unei enzime autolitice care degradeaz
cortexul sporal. Are loc absorbia de ap, eliberarea dipicolinatului
de calciu i degradarea unor compui sporali.Dezvoltarea sporului.
Protoplastul sporal se transform n bacterie vegetativ, care trece
printr-o perioad metabolic activ de refacere a constituenilor
celulari normali i a echipamentului enzimatic complet.