Embed Size (px)
DESCRIPTION
十一章 细胞质 遗传. 第一 节细胞质遗传的概念和特点 细胞质遗传:由细胞质内的基因即细胞质基因所决定的遗传现象和遗传规律称之。 非染色体遗传,非孟德尔遗传,核外遗传,母体遗传,母性遗传. 特点:. 遗传方式非孟德尔式:杂交后代不表现一定比例的分离。 正交和反交的遗传表现不同。 通过连续回交能将母本的核基因几乎全部置换。 由附加体或共生体决定的性状,其表现往往类似病毒的转导或感染。. 第三节 叶绿体遗传. 1 ) 1908 , Carl Corrans ,非孟德尔遗传发现者,紫茉莉花斑枝条. (一) 叶绿体系统的核外遗传. - PowerPoint PPT Presentation
Citation preview

十一章 细胞质十一章 细胞质遗传遗传第一节细胞质遗传的概念和特点第一节细胞质遗传的概念和特点
•细胞质遗传:由细胞质内的基因即细细胞质遗传:由细胞质内的基因即细胞质基因所决定的遗传现象和遗传规胞质基因所决定的遗传现象和遗传规律称之。律称之。
•非染色体遗传,非孟德尔遗传,核外非染色体遗传,非孟德尔遗传,核外遗传,母体遗传,母性遗传遗传,母体遗传,母性遗传

特点:特点:
• 遗传方式非孟德尔式:杂交后代不表现一遗传方式非孟德尔式:杂交后代不表现一定比例的分离。定比例的分离。
• 正交和反交的遗传表现不同。正交和反交的遗传表现不同。• 通过连续回交能将母本的核基因几乎全部通过连续回交能将母本的核基因几乎全部
置换。置换。• 由附加体或共生体决定的性状,其表现往由附加体或共生体决定的性状,其表现往
往类似病毒的转导或感染。往类似病毒的转导或感染。

第三节 叶绿体遗传第三节 叶绿体遗传

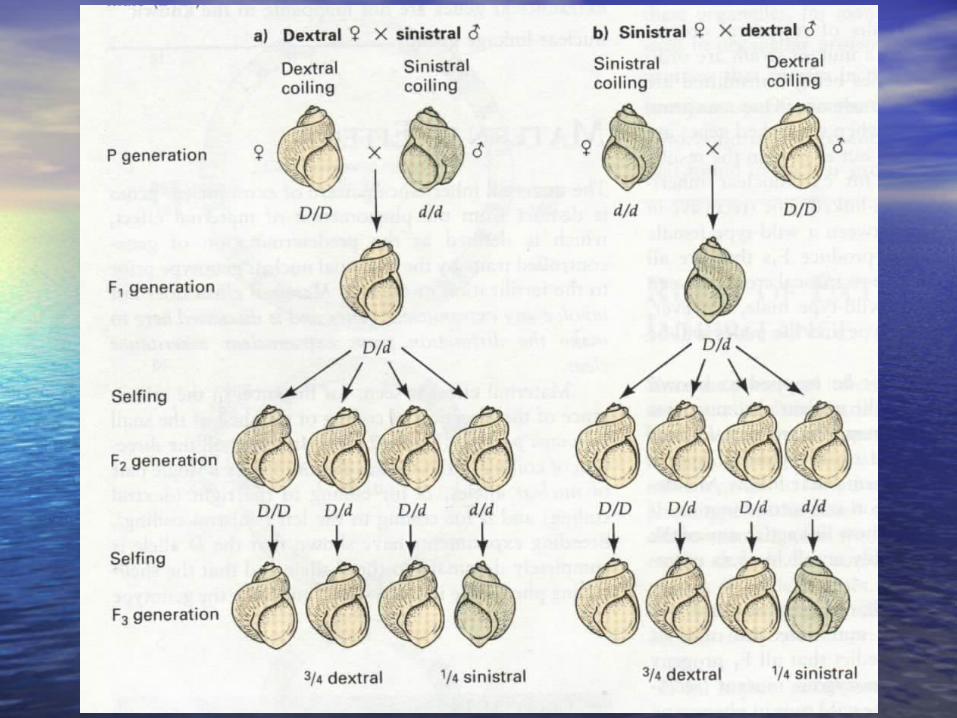
11 )) 19081908 ,, Carl CorransCarl Corrans ,非孟德尔遗传发,非孟德尔遗传发现者,紫茉莉花斑枝条现者,紫茉莉花斑枝条


(一)(一)叶绿体系统的核外遗传叶绿体系统的核外遗传• 19091909 年由年由 Carl CorransCarl Corrans 发现(发现( irabilis jalapairabilis jalapa ))
表 20-8 紫茉莉花斑植物杂交的结果母本枝条表型 父本枝条表型 后代表型
白色 白色绿色 白色
白色
花斑 白色白色 绿色绿色 绿色
绿色
花斑 绿色白色 花斑,绿色,白色绿色 花斑,绿色,白色
花斑
花斑 花斑,绿色,白色

叶绿体叶绿体 DNA—— ct DNADNA—— ct DNA
• 环状双链结构环状双链结构
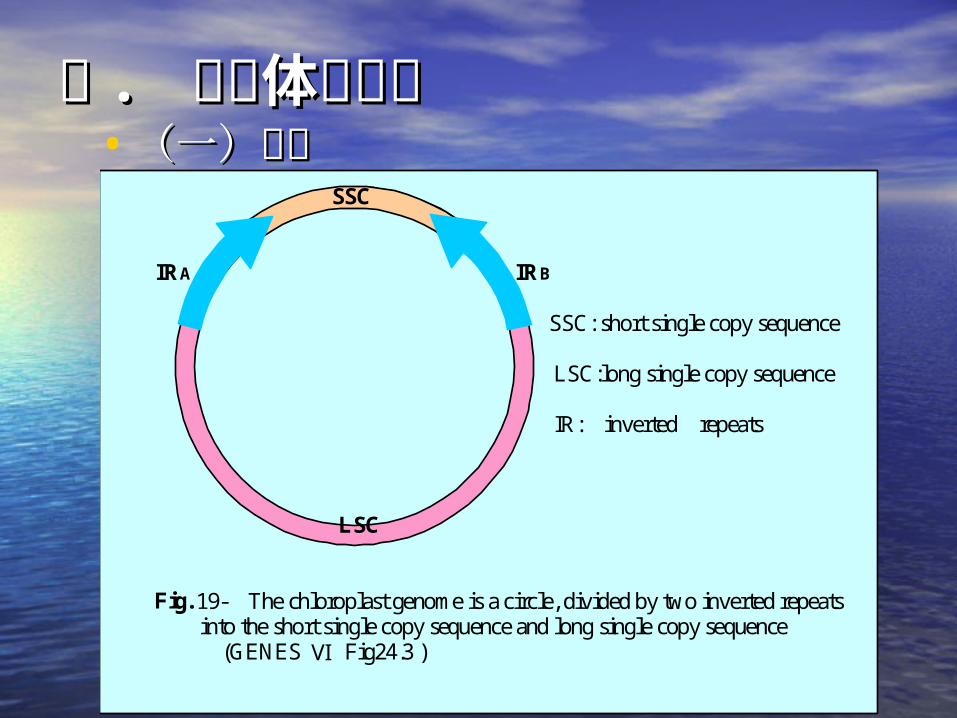
二二 . . 叶绿体基因组叶绿体基因组• (一)结构(一)结构 SSC
IRA IRB
SSC: short single copy sequence LSC:long single copy sequence IR: inverted repeats
LSC
Fig. 19- The chloroplast genome is a circle, divided by two inverted repeats into the short single copy sequence and long single copy sequence (GENES Ⅵ Fig24.3 )
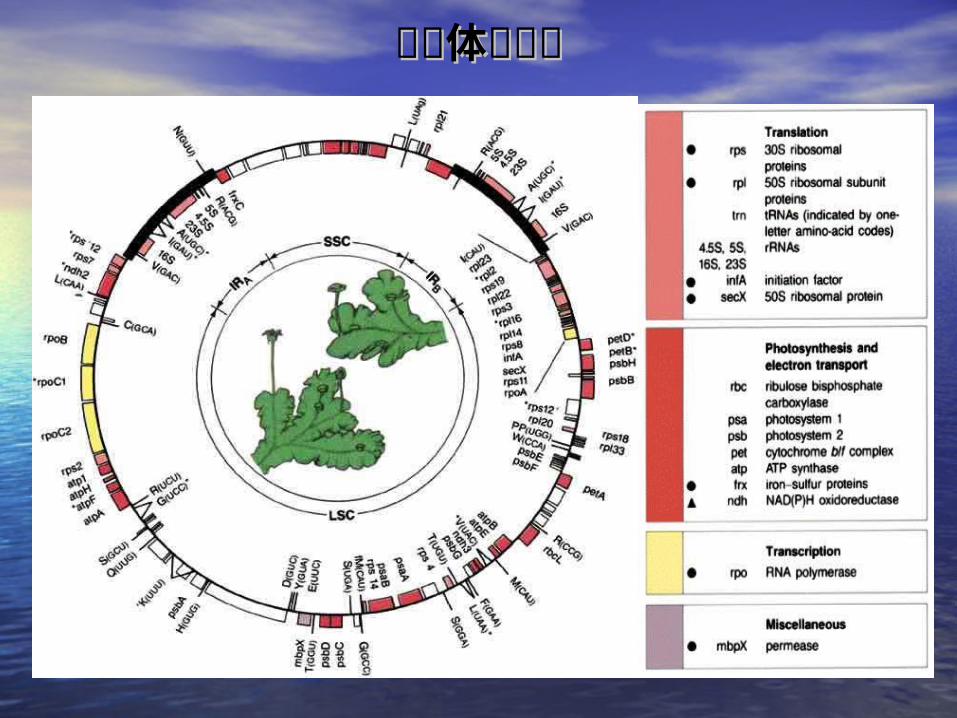
叶绿体基因组叶绿体基因组

Ct DNACt DNA 的基因组成的基因组成
表 20-6叶绿体基因组的组成 地钱 烟草 水稻
反向重复序列 10,058 25,339 20,798短单拷贝序列 19,813 18,482 12,335长单拷贝序列 81,095 86,684 80,592总基因组 121,024 155,844 134,525参考 B.Lewi n: 《GENES Ⅵ》 . 1997, Tabl e 24。3

Ct DNACt DNA 基因组成有以下特点:基因组成有以下特点:(( 11 )基因组由两个)基因组由两个 IRIR 和一个 和一个 SSCSSC 及一个 及一个 LSLS(( 22 )) IRAIRA 和和 IRBIRB ,编码相同,方向相反。,编码相同,方向相反。(( 33 )) ctDNActDNA 启动子和原核生物的相似,基因启动子和原核生物的相似,基因产生单顺反子或多顺反子的产生单顺反子或多顺反子的 mRNAmRNA ;;
(( 44 )不同)不同 ctDNActDNA 基因组成和数目几乎是相同基因组成和数目几乎是相同的,产物多为类囊体的成分或和氧化还原反应的,产物多为类囊体的成分或和氧化还原反应有关;有关;
(( 55 )其)其 tRNAtRNA 基因中有内含子,有的位于基因中有内含子,有的位于 DD 环环上, 此和原核及真核生物核上, 此和原核及真核生物核 tRNAtRNA 都不相同;都不相同;
(( 66 )所有叶绿体基因转录的)所有叶绿体基因转录的 mRNAmRNA 都由叶绿都由叶绿体核糖体翻译。体核糖体翻译。
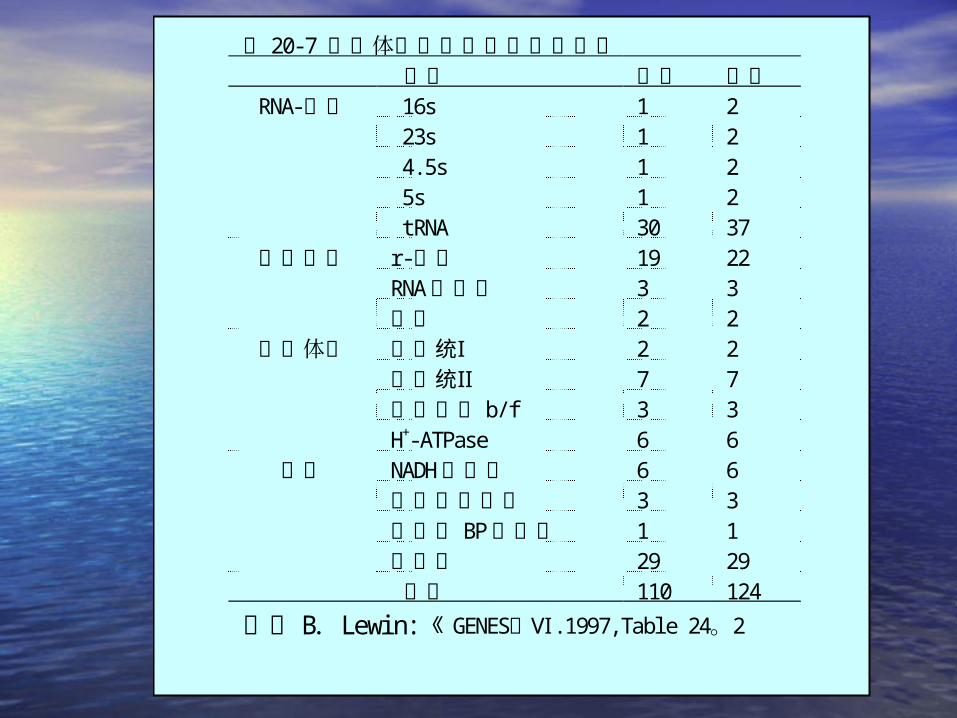
表 20-7 叶绿体基因组编码的各种蛋白基因 类型 拷贝
16s 1 2 23s 1 2 4. 5s 1 2 5s 1 2
RNA-编码
tRNA 30 37r-蛋白 19 22RNA聚合酶 3 3
基因表达
其它 2 2Ⅰ光系统 2 2Ⅱ光系统 7 7
细胞色素 b/ f 3 3
类囊体膜
H+-ATPase 6 6NADH脱氢酶 6 6铁氧化还原酶 3 3核酮糖 BP羧化酶 1 1
其它
未鉴定 29 29总计 110 124
参考 B.Lewi n: 《GENES Ⅵ》 . 1997, Tabl e 24。2

第四节 第四节 线粒体线粒体遗传遗传
一一 .. 线粒体基因组线粒体基因组 (mt DNA)(mt DNA)((一)一) mt DNAmt DNA 的基因组成的基因组成 (1)(1) 闭合环状闭合环状 DNADNA (2) (2) 基因数目和排列顺序相同基因数目和排列顺序相同(( 33 )有)有 DD 环和环和 22 个复制起始点个复制起始点(( 55 )基因间没有间隔,因此每个基因不可)基因间没有间隔,因此每个基因不可 能都有自己的起动子能都有自己的起动子
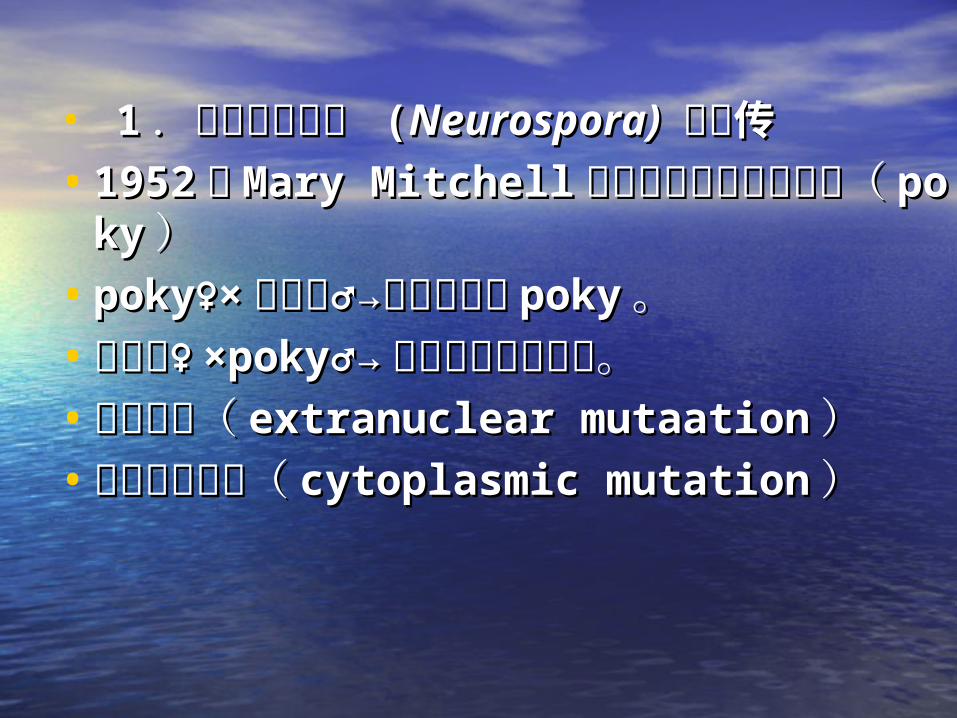
• 11 .小菌落脉孢菌 .小菌落脉孢菌 ((Neurospora) Neurospora) 的遗传的遗传• 19521952 年年 Mary MitchellMary Mitchell 分离了脉胞菌的小菌分离了脉胞菌的小菌
落(落( pokypoky ))• poky♀×poky♀× 野生型♂→后代全部为野生型♂→后代全部为 pokypoky 。。• 野生型♀野生型♀ ×poky♂→×poky♂→ 后代全部为野生型。后代全部为野生型。• 核外突变(核外突变( extranuclear mutaationextranuclear mutaation ))• 或细胞质突变(或细胞质突变( cytoplasmic mutationcytoplasmic mutation ))

酵母小菌落酵母小菌落 (peptite)(peptite) 突变突变
• 19401940 ,, Boris EphrnssiBoris Ephrnssi• 在正常的细胞群体中有在正常的细胞群体中有 0.1%0.1% ~~ 1%1%
的细胞会自发变成小菌落。在插入剂的细胞会自发变成小菌落。在插入剂(如:溴化乙锭,(如:溴化乙锭, EBEB )存在时)存在时 100100%% 的细胞都变成小菌落。的细胞都变成小菌落。
• 分类 分类 (1) (1) 核基因突变型核基因突变型-分离型-分离型 (2) (2) 中性型中性型-- mtDNAmtDNA 丢失丢失 (3) (3) 抑制型抑制型- - mtDNAmtDNA 突变突变

酵母小菌落酵母小菌落
表 20-9酵母小菌落的类型和特点 小菌落 类型
符号 产生 原因
与野生型 杂交
二倍体表型 四分体的基因型, 表型和分离比
分离型 pet - 核 基因 突变
pet -× pet+ → pet -/ pet+ (正常)
→小菌落 (pet -) :大菌落 =2: 2
中性型 [P -N] MtDNA丢失
[P -N]× [P +N] → [P -N] / [P +N] → (正常)
→全为大菌落[P +
N] 抑制型 [P -S] MtDNA
突变 [P -S]× [P +S] → [P -S] / [P +S] →
(部分呼吸) →小菌落[P -S] (迅速形成孢子) →小菌落[P -S] (长时间分裂) 比例不等 (1~99%)

正常线粒体 小菌落线粒体正常线粒体 小菌落线粒体
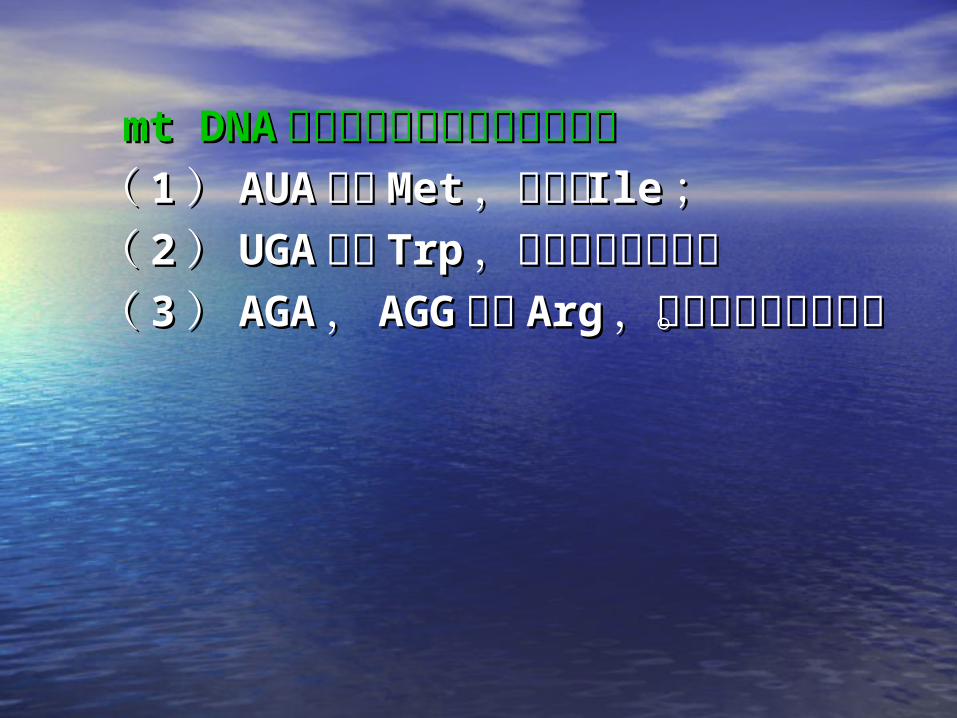
mt DNAmt DNA 密码子与核基因密码子的差异密码子与核基因密码子的差异(( 11 )) AUAAUA 编码编码 MetMet ,而不是,而不是 IleIle ;;(( 22 )) UGAUGA 编码编码 TrpTrp ,而不是终止密码,而不是终止密码
子子(( 33 )) AGAAGA ,, AGGAGG 编码编码 ArgArg ,而不是终,而不是终
止密码子。止密码子。

表 20-2 在线粒体中密码子的改变
生物 密码子 MtDNA编码 核密码 共同 UGA Trp 终止 哺乳动物 AGA/ AGG 终止 Arg 哺乳动物 AUA Met(起始) I l e 果蝇 AUA Met I l e 果蝇 AGA Ser Arg 纤毛虫 UAA/ UAG Gl u 终止 另一类纤毛虫 UGA/ UAG Cys 终止 酵母 AUA Met(延伸) I l e 酵母 CCA Thr Leu 酵母 CUG Ser Leu 酵母 UAG Ser 终止 支原体 UGA Trp 终止
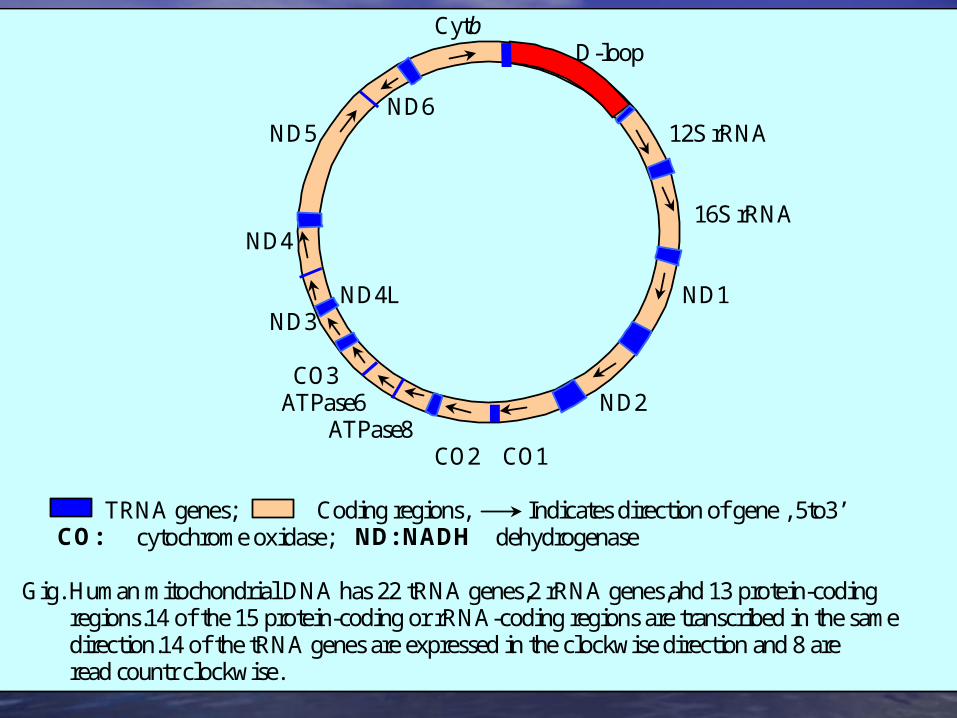
Cytb D-loop ND6 ND5 12S rRNA
16S rRNA ND4 ND4L ND1 ND3 CO3 ATPase6 ND2 ATPase8 CO2 CO1 TRNA genes; Coding regions, Indicates direction of gene , 5to3’ CO: cytochrome oxidase; ND: NADH dehydrogenase Gig. Human mitochondrial DNA has 22 tRNA genes,2 rRNA genes,ahd 13 protein-coding regions.14 of the 15 protein-coding or rRNA-coding regions are transcribed in the same direction.14 of the tRNA genes are expressed in the clockwise direction and 8 are read countr clockwise.

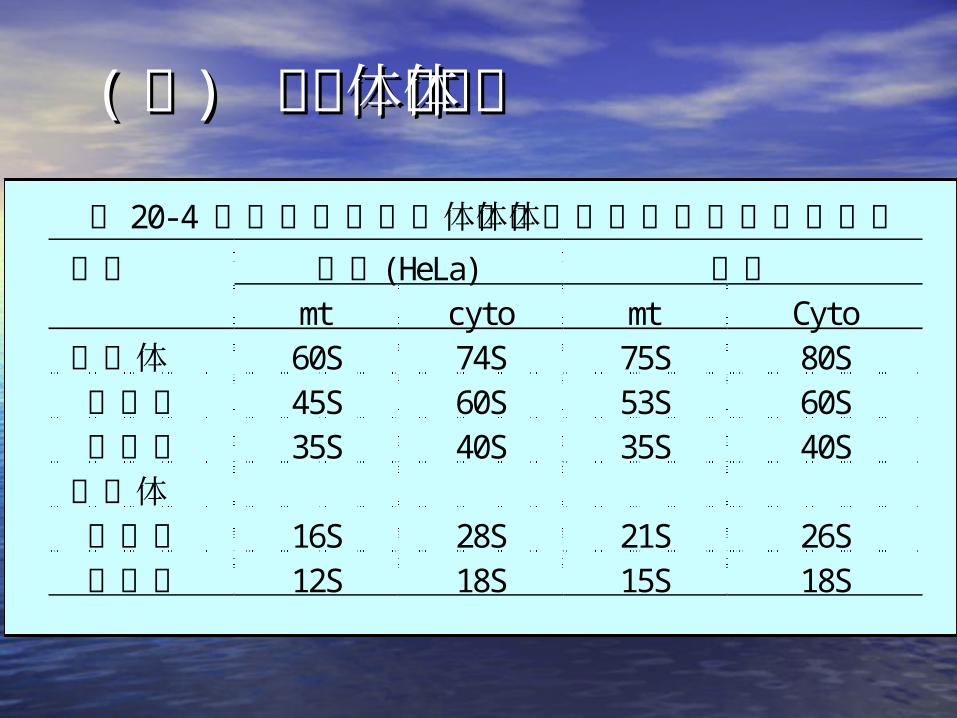
(( 二二 ) ) 线粒体核糖体线粒体核糖体 表 20-4 人类和酵母线粒体核糖体和细胞质核糖体的大小
人类(HeLa) 酵母成分mt cyto mt Cyto
核糖体 60S 74S 75S 80S 大亚基 45S 60S 53S 60S 小亚基 35S 40S 35S 40S核糖体 大亚基 16S 28S 21S 26S 小亚基 12S 18S 15S 18S
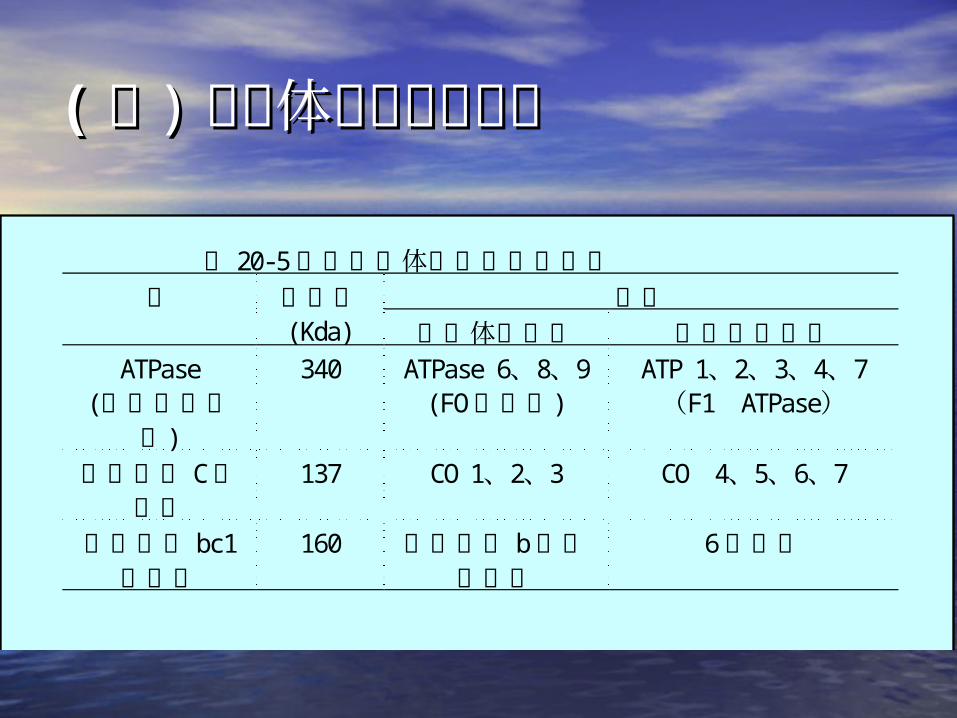
(( 三三 )) 线粒体蛋白质的合成线粒体蛋白质的合成
表 20-5酵母线粒体中合成的酶亚基亚基酶 分子量
(Kda) 线粒体中合成 细胞质中合成ATPase
(对寡霉素敏感)
340 ATPase 6、8、9(FO膜因子)
ATP 1、2、3、4、7(F1 ATPase)
细胞色素 C氧化酶
137 CO 1、2、3 CO 4、5、6、7
细胞色素 bc1复合物
160 细胞色素 b脱辅基蛋白
6个亚基
线粒体的复制线粒体的复制

线粒体的起源线粒体的起源
• 内共生假说内共生假说• 19701970 ,, MargulisMargulis ,真核,真核
细胞祖先是种吞噬细胞;细胞祖先是种吞噬细胞;线粒体祖先是种革兰氏阴线粒体祖先是种革兰氏阴性菌。前者吞后者性菌。前者吞后者
• 细胞分化假说细胞分化假说• 原始的原核细胞质膜内陷原始的原核细胞质膜内陷
包被包被 DNADNA ,然后再分化,然后再分化形成独立的细胞器形成独立的细胞器。。
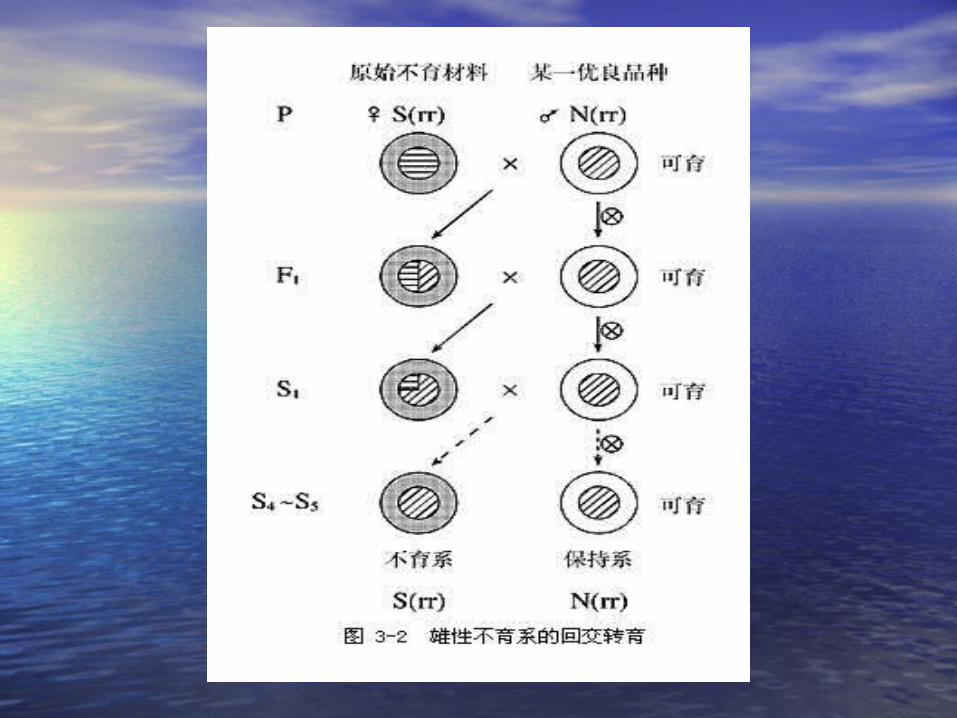
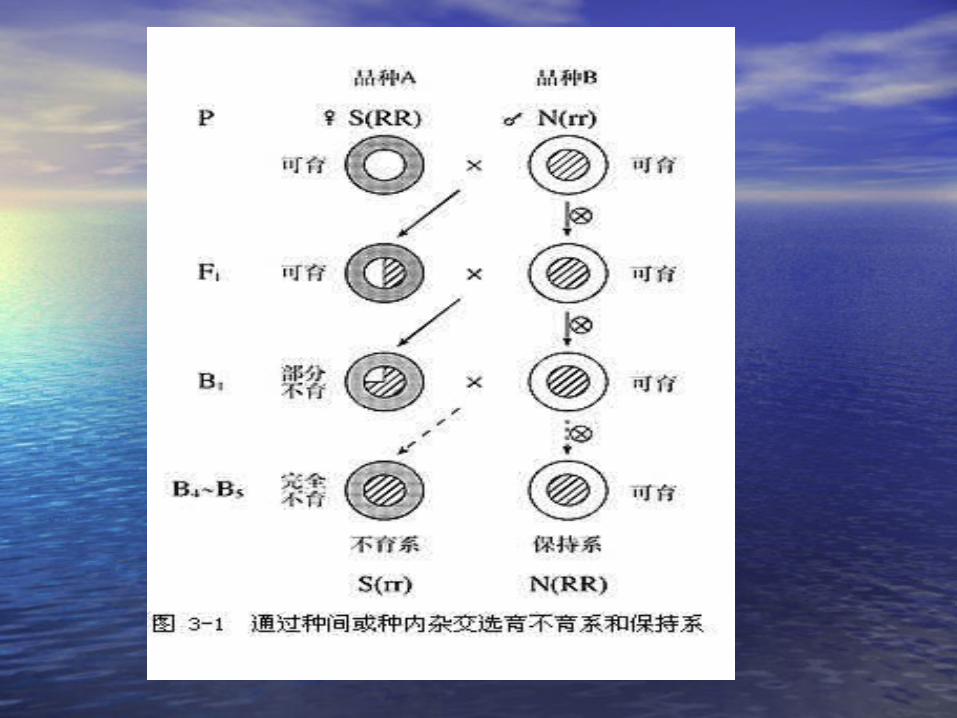
第六节 植物雄性不育的遗传第六节 植物雄性不育的遗传
• 雄性不育(雄性不育( male sterility) male sterility) 的类别及其遗的类别及其遗传特点传特点
• 核不育型:由核内染色体上的基因所决定的核不育型:由核内染色体上的基因所决定的雄性不育类型。雄性不育类型。
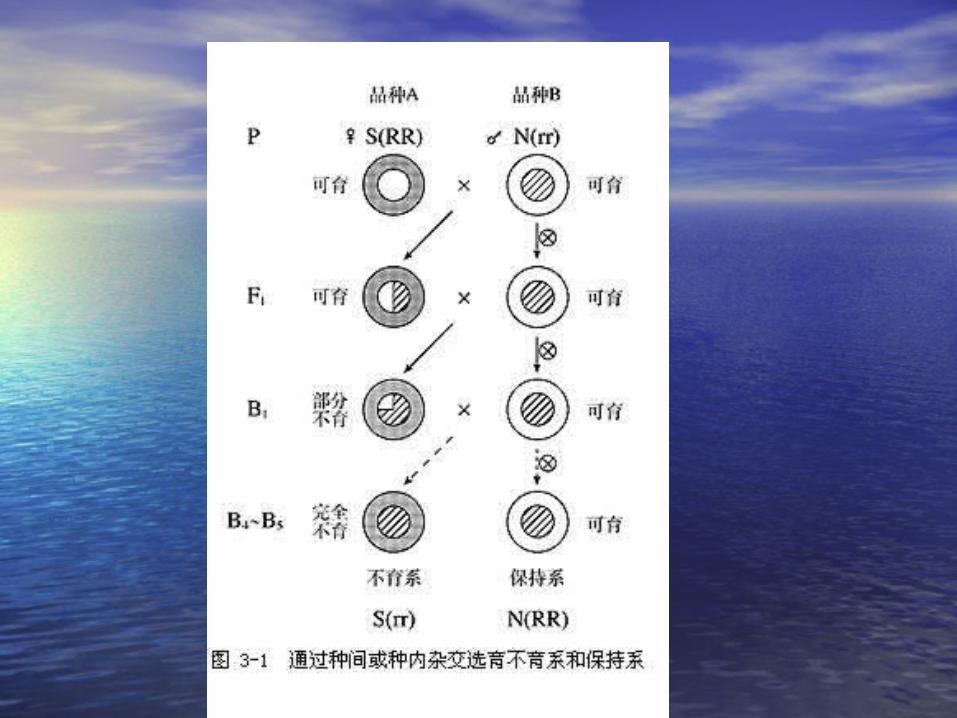
• 质核不育型:由细胞质基因和核基因互作控质核不育型:由细胞质基因和核基因互作控制的不育类型(制的不育类型( CMSCMS ) )

• 细胞质中与雄性不育有关的基因:细胞质中与雄性不育有关的基因: S-S- 不育不育 N-N- 可育可育• 细胞核中与雄性不育有关的基因:细胞核中与雄性不育有关的基因: R-rR-r• RR Rr-RR Rr- 可育 可育 rr –rr – 不育不育
S(rr)S(rr) ,, S(RR),S(RR), S(Rr) S(Rr) N(RR),N(RR), N(Rr), N(Rr), N(rr)N(rr)
不育系
保持系
恢复系
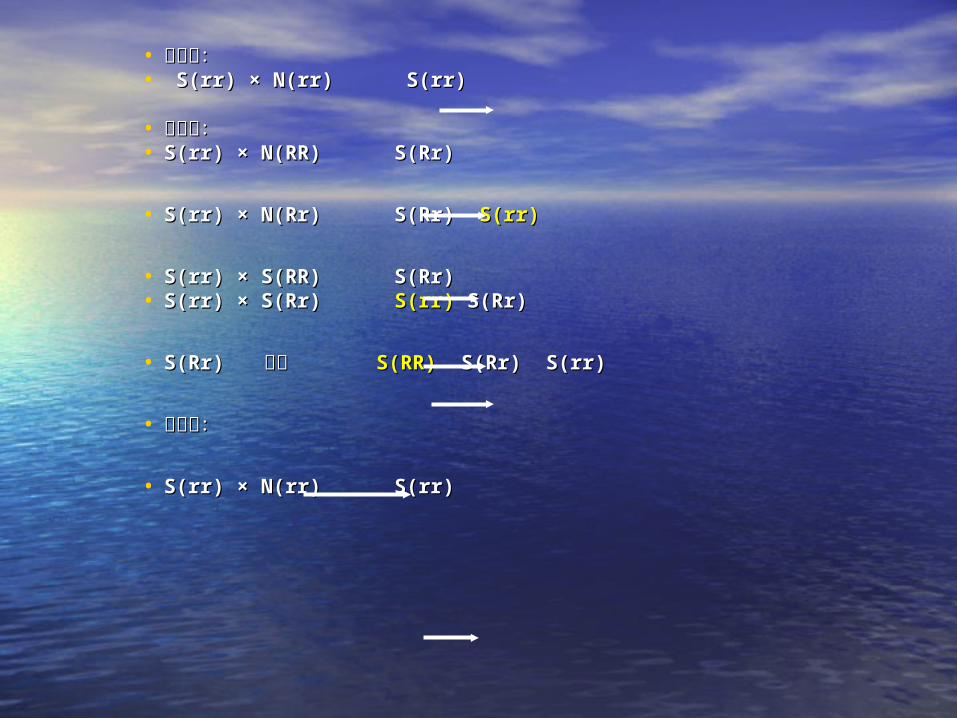
• 不育系:不育系:• S(rr) × N(rr) S(rr)S(rr) × N(rr) S(rr)
• 恢复系:恢复系:• S(rr) × N(RR) S(Rr)S(rr) × N(RR) S(Rr)
• S(rr) × N(Rr) S(Rr) S(rr) × N(Rr) S(Rr) S(rr)S(rr)
• S(rr) × S(RR) S(Rr)S(rr) × S(RR) S(Rr)• S(rr) × S(Rr) S(rr) × S(Rr) S(rr)S(rr) S(Rr) S(Rr)
• S(Rr) S(Rr) 自交 自交 S(RR)S(RR) S(Rr) S(rr) S(Rr) S(rr)
• 保持系:保持系:
• S(rr) × N(rr) S(rr)S(rr) × N(rr) S(rr)

第六节 植物雄性不育的遗传第六节 植物雄性不育的遗传
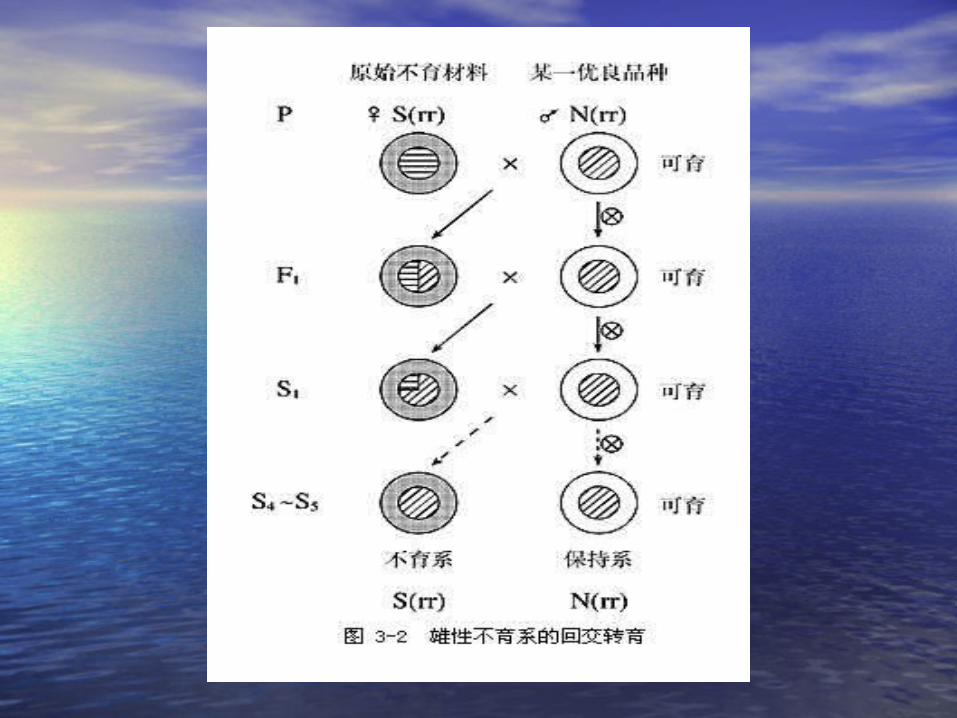
•三系制种法三系制种法。。• (1)(1) 雄性不育系:雄性不育系: S(rr)S(rr) 甲甲♀♀×♂×♂ 乙组合中将甲转成不育系乙组合中将甲转成不育系•(( 22 )保持系:)保持系: NN (( rr) rr) •(( 33 )恢复系:杂交组合中的父本必须是恢)恢复系:杂交组合中的父本必须是恢复系。复系。 N(RR) , S(RR)N(RR) , S(RR)
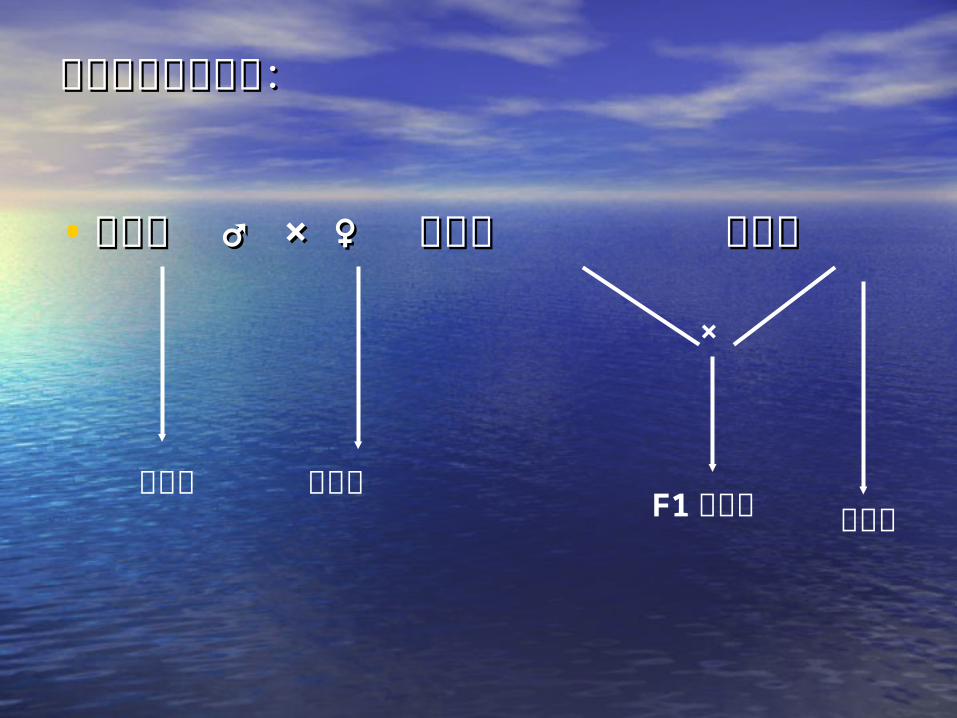
三系法制种示意图:三系法制种示意图:
• 保持系 保持系 ♂♂ × × ♀♀ 不育系 恢复系不育系 恢复系
×
保持系 不育系 F1 杂交种 恢复系

水稻专家——袁隆平水稻专家——袁隆平

在生产上配制杂交种应注意的问题在生产上配制杂交种应注意的问题 • 关于保持系:必须具备两个条件关于保持系:必须具备两个条件• 11、用它作父本,与不育系杂交所产生的后代始、用它作父本,与不育系杂交所产生的后代始
终是不育的;终是不育的;• 22、除了它本身可育外,其他性状应当与不育系、除了它本身可育外,其他性状应当与不育系完全相同。完全相同。
• 关于恢复系 关于恢复系 • 如某优良杂交组合的父本无恢复能力,即核的育如某优良杂交组合的父本无恢复能力,即核的育
性基因为性基因为 rrrr ,就需将恢复基因,就需将恢复基因 RR 转入到该父本中。转入到该父本中。• 用不育系和恢复系生产杂交种时,为了使天然受用不育系和恢复系生产杂交种时,为了使天然受粉能顺利地进行,不育系与恢复系的花期必须一粉能顺利地进行,不育系与恢复系的花期必须一致,或者不育系的开花稍早一些。一般恢复系的致,或者不育系的开花稍早一些。一般恢复系的植株应比不育系的植株高,以便花粉能很好地散植株应比不育系的植株高,以便花粉能很好地散落到不育系的花上 。落到不育系的花上 。

建立两个隔离区建立两个隔离区
•繁殖不育系和保持系的隔离区,繁殖不育系和保持系的隔离区,• 制杂交种的隔离区。 制杂交种的隔离区。
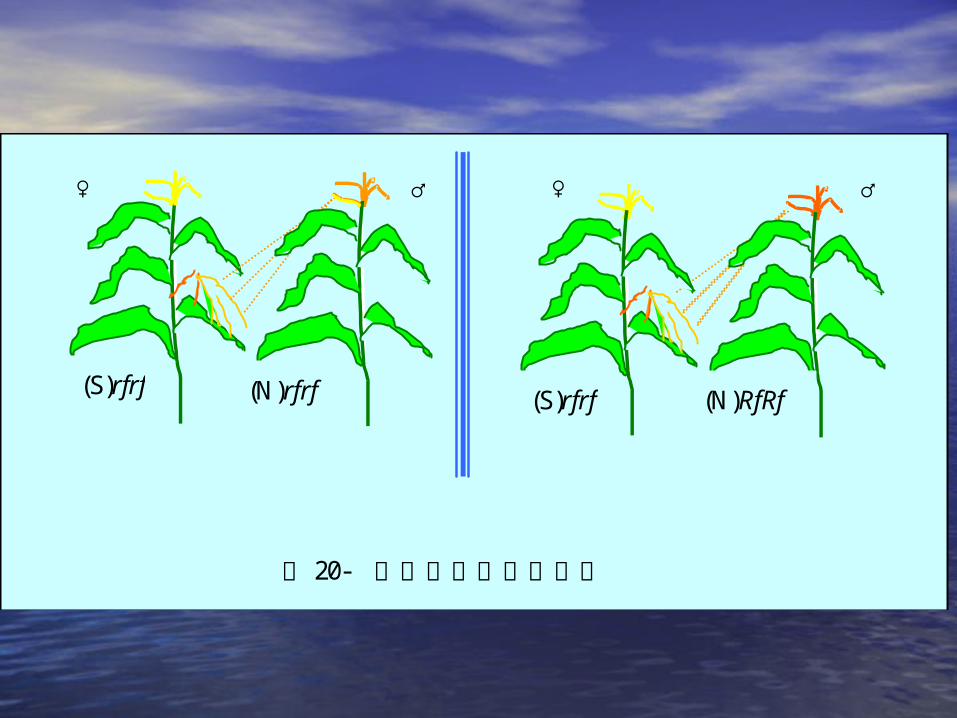
♀ ♂ ♀ ♂
图 20- 玉米的三系两区育种
(S)rfrf (N)rfrf (S)rfrf (N)RfRf

二系法制种二系法制种
•石明松:石明松: 19731973 年从农垦年从农垦 5858(晚粳稻)(晚粳稻)中发现光敏核不育水稻——农垦中发现光敏核不育水稻——农垦 58S58S
• 特点:特点:•长日照——不育长日照——不育•短日照——可育短日照——可育
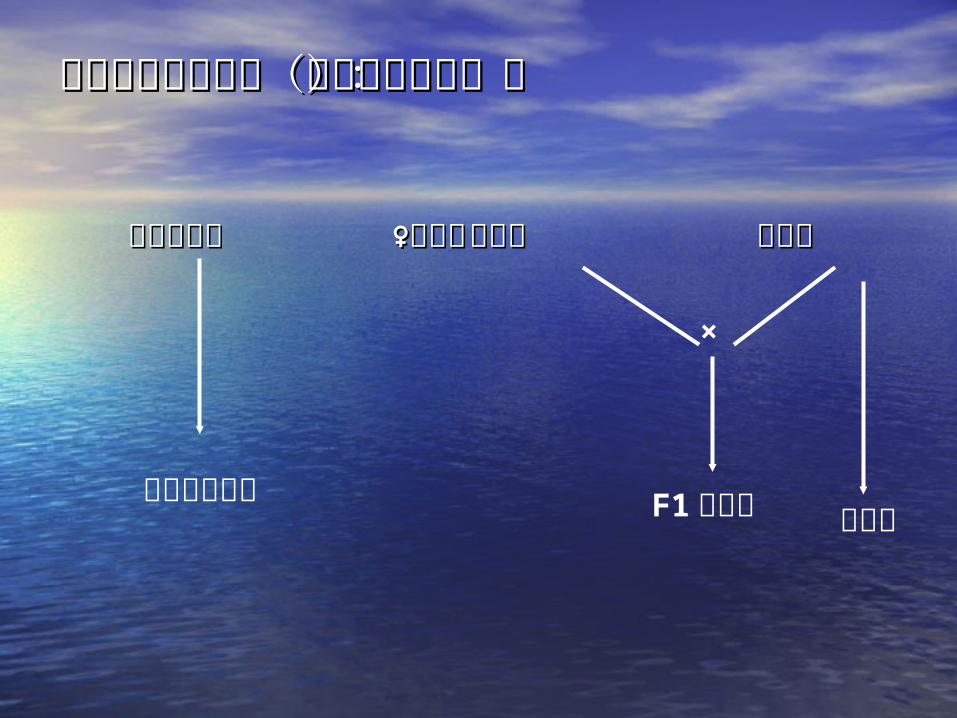
二系法制种示意图(光敏核不育系):二系法制种示意图(光敏核不育系):
短日照可育短日照可育 ♀长日照♀长日照不育系不育系 恢复系恢复系
×
光敏核不育系 F1 杂交种 恢复系

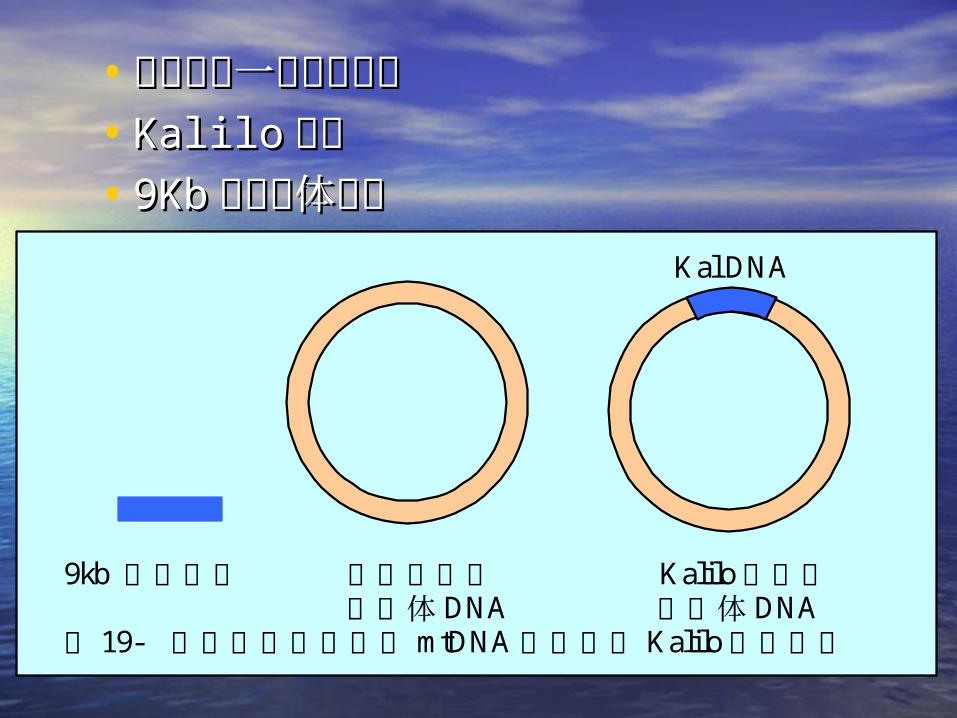
• 脉胞菌中一种衰老品系脉胞菌中一种衰老品系• KaliloKalilo 品系品系• 9Kb9Kb 的线粒体质粒的线粒体质粒
Kal DNA
9kb 线性质粒 正常品系的 Kalilo品系的 线粒体 DNA 线粒体 DNA 图 19- 脉孢菌衰老品系的 mtDNA中插入了 Kalilo线性质粒

第三节 母体影响第三节 母体影响(( maternal effecmaternal effectt ))
•母体的基因型决定后代表型的现象称为母体的基因型决定后代表型的现象称为母体影响母体影响(( maternal effectmaternal effect ))
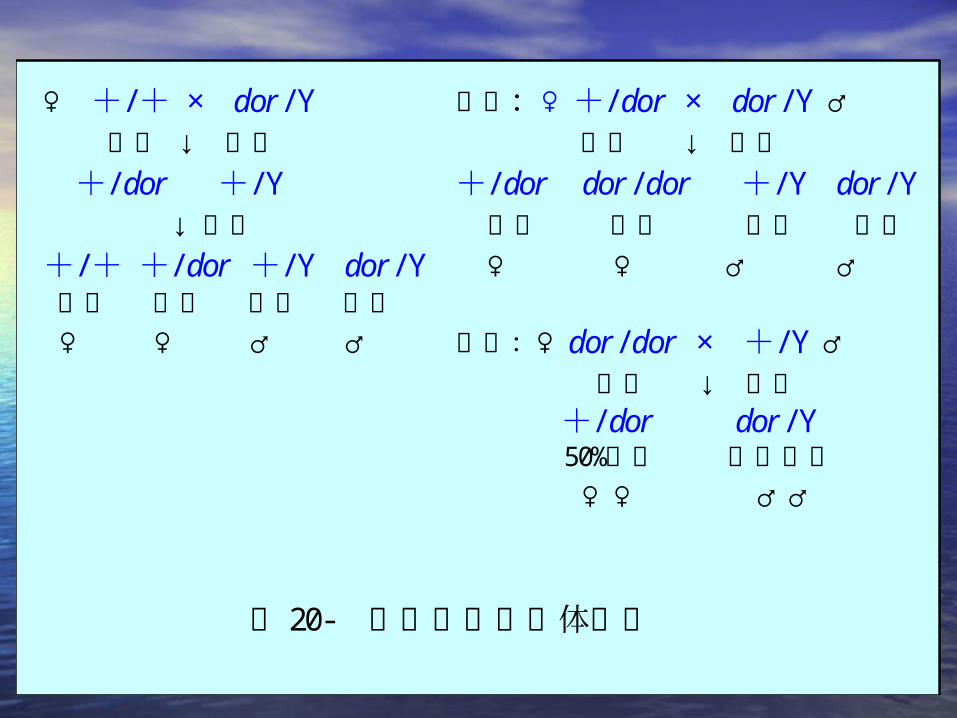
♀ +/+ × dor/ Y 回交:♀+/ dor × dor/ Y ♂ 红眼 ↓ 黄眼 红眼 ↓ 黄眼 +/ dor +/ Y +/ dor dor/dor +/ Y dor/ Y ↓互交 红眼 黄眼 红眼 黄眼+/+ +/ dor +/ Y dor/ Y ♀ ♀ ♂ ♂ 红眼 红眼 红眼 黄眼 ♀ ♀ ♂ ♂ 反交:♀ dor/dor × +/ Y ♂ 黄眼 ↓ 红眼 +/ dor dor/ Y 50%致死 胚胎致死 ♀ ♀ ♂♂
图 20- 果蝇眼色的母体影响